


Abstracto
Los ortoflavivirus comprenden un género diverso de virus de ARN de cadena positiva que incluye importantes patógenos humanos como el dengue, la fiebre amarilla, el Zika, la encefalitis japonesa, la encefalitis transmitida por garrapatas y el virus del Nilo Occidental. A pesar de su impacto global, las restricciones moleculares que preservan la replicación viral en distintos vectores y entornos de hospedadores aún no se comprenden completamente. La replicación del ARN viral depende del bucle tallo A (SLA), un elemento de ARN estructurado ubicado en el extremo 5′ del genoma que recluta la polimerasa viral NS5. En este estudio, examinamos la diversidad estructural de este promotor de ARN en los ortoflavivirus. Mediante clones infecciosos, virus reporteros y análisis estructurales computacionales, demostramos que los SLA de diversos virus transmitidos por mosquitos y garrapatas son funcionalmente intercambiables en el contexto de clones infecciosos de los virus del dengue y del Zika. Los análisis de estructura-función revelan que los contactos de nucleótidos conservados entre el bucle superior del SLA y el dominio de la polimerasa NS5 forman una interfaz de interacción conservada que se mantiene en todos los ortoflavivirus, incluidos los virus específicos de insectos. En contraste, otros subelementos de SLA, como la unión de tres vías y el tallo lateral, han divergido de manera específica para cada grupo; sin embargo, la coevolución en virus transmitidos por mosquitos y garrapatas conserva la arquitectura tridimensional y la capacidad de unión a NS5 para la replicación viral. Guiados por esta conservación, identificamos pequeñas moléculas que se unen a SLA e inhiben la replicación en múltiples ortoflavivirus patógenos. Estos hallazgos revelan principios fundamentales que rigen la evolución del promotor de ARN viral y establecen estructuras de ARN conservadas como objetivos prometedores para el control viral.
Resumen del autor
Figuras
Cita: Oviedo-Rouco S, Bertoni L, Mikkelsen E, Sarto C, Gonzalez Lopez Ledesma MM, Pallarés HM, et al. (2026) El promotor de ARN para la replicación de ortoflavivirus patógenos es universal y sirve como diana para la inhibición viral. PLoS Pathog 22(5): e1014233. https://doi.org/10.1371/journal.ppat.1014233
Editor: Peter Sarnow, Universidad de Stanford, ESTADOS UNIDOS DE AMÉRICA
Recibido: 10 de marzo de 2026; Aceptado: 4 de mayo de 2026; Publicado: 18 de mayo de 2026
Copyright: © 2026 Oviedo-Rouco et al. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución , que permite el uso, la distribución y la reproducción sin restricciones en cualquier medio, siempre que se cite al autor original y la fuente.
Disponibilidad de datos: Los datos presentados en este manuscrito están disponibles en el artículo y en su material complementario.
Financiación: SOR, LB, CS y HMP recibieron becas del CONICET. Este trabajo fue financiado por PICT-2019-02869 y el Programa Conjunto de Investigación en Salud Canadá-Israel, CIHR/IRSC/ISF (495147) para AVG. También se obtuvo financiación de la Universidad de Duke y la Universidad de Toronto Mississauga para AH. Los financiadores no participaron en el diseño del estudio, la recopilación e interpretación de datos ni en la decisión de enviar el trabajo para su publicación.
Conflictos de intereses: He leído la política de la revista y los autores de este manuscrito tienen los siguientes conflictos de intereses: AVG, MMGLL, CVF y MA son miembros del Consejo Nacional de Investigación y Desarrollo (CONICET) de Argentina.
Introducción
Los ortoflavivirus son un grupo diverso de virus de ARN monocatenario de sentido positivo y envueltos que incluyen importantes patógenos humanos emergentes y reemergentes. Entre ellos, el virus del dengue (DENV), el virus del Nilo Occidental (WNV), el virus del Zika (ZIKV), el virus de la encefalitis transmitida por garrapatas (TBEV) y el virus de la fiebre amarilla (YFV) impulsan epidemias recurrentes con morbilidad y mortalidad sustanciales [ 1 , 2 ]. Es de gran preocupación que se hayan notificado brotes de otros ortoflavivirus menos caracterizados en humanos y animales en diferentes regiones del mundo [ 3 ]. El dengue, la enfermedad viral transmitida por artrópodos más prevalente, infecta a unos 400 millones de humanos cada año; más de una cuarta parte de la población mundial vive en áreas donde el DENV es ahora endémico [ 4 ]. En 2024, epidemias sin precedentes de DENV azotaron las Américas, con brotes que se extendieron a países sin antecedentes de dengue [ 5 ]. El ZIKV mostró su potencial epidémico durante los brotes de 2015-2017 en las Américas, cuando la infección se relacionó con malformaciones congénitas, incluida la microcefalia, y complicaciones neurológicas [ 6 ]. El YFV se asocia con una alta mortalidad y ha resurgido en Sudamérica a pesar de la disponibilidad de una vacuna eficaz [ 7 ].
Estos virus infectan una amplia gama de huéspedes vertebrados e invertebrados y se clasifican ecológicamente según su ciclo de transmisión en transmitidos por mosquitos (MBFV), transmitidos por garrapatas (TBFV), específicos de insectos (ISFV) y flavivirus sin vector conocido (NKVFV) ( Fig. 1A ). Los virus asociados con enfermedades humanas se transmiten predominantemente por mosquitos o garrapatas. Se han dedicado esfuerzos considerables al desarrollo de vacunas contra varios miembros del género, lo que ha llevado a la aprobación y distribución de vacunas contra YFV, virus de la encefalitis japonesa (JEV), TBEV y, más recientemente, DENV [ 8 – 12 ]. Se espera que estas vacunas jueguen un papel clave en el control de enfermedades.
-
PNGimagen más grande
-
PELEAimagen original
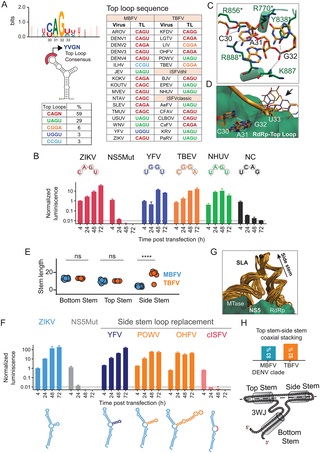
A. Árbol filogenético de ortoflavivirus construido a partir de un alineamiento de secuencias múltiples de la secuencia NS5. Los diferentes grupos ecológicos se indican con diferentes colores. B. Representación esquemática del mecanismo de iniciación de la síntesis de ARN de cadena negativa del DENV [ 31 ]. Se indican el SLA y el 3’SL. Se muestran las formas lineal y circular del genoma. El dominio RdRp de NS5 inicia la síntesis de ARN en el extremo 3′ del genoma circularizado. C. Representación esquemática del complejo de ribonucleoproteína SLA-NS5 determinado por crio-EM, que muestra los dos puntos de contacto: el bucle superior del SLA con el RDRP y el extremo 5′ del ARN con la MTasa de NS5. SLA en naranja y NS5 en verde. ID del banco de datos de proteínas 8gzp. D. Modelos 2D y 3D para SLA seleccionados correspondientes a MBFV, TBFV e ISFV, como se indica a la derecha. Se muestran dos modelos para cada secuencia en cintas opacas y translúcidas para resaltar la variabilidad entre las conformaciones SLA. E. Conservación de nucleótidos en función de la posición calculada a partir del alineamiento de secuencias SLA. F. Representación gráfica de la conservación estructural de cada nucleótido a partir de un alineamiento secuencia-estructura. Se indica el código de color de 0 a 100 %. G. Logotipo de secuencia que muestra la conservación de nucleótidos obtenida del alineamiento secuencia-estructura SLA construido utilizando secuencias de cada ortoflavivirus disponible. Para mayor claridad, la numeración se ajustó al genoma del DENV. Los dibujos utilizados en esta figura fueron de imágenes prediseñadas de dominio público tomadas de NIH Bio Art. https://bioart.niaid.nih.gov/bioart/162 , https://bioart.niaid.nih.gov/bioart/11 , https://bioart.niaid.nih.gov/bioart/247 , https://bioart.niaid.nih.gov/bioart/49 , persona de pie modificado de: https://bioart.niaid.nih.gov/bioart/555 .
Paralelamente, la extensa investigación sobre descubrimiento de fármacos se ha centrado en las proteínas virales. A pesar de estos esfuerzos, no se ha aprobado ningún fármaco antiviral específico para el tratamiento de ninguna infección por ortoflavivirus [ 13 , 14 ]. Esta brecha ha impulsado un creciente interés en nuevas estrategias terapéuticas, incluyendo ARN virales relevantes como objetivos de intervención. Aunque los estudios de prueba de concepto demuestran que las estructuras de ARN pueden ser objetivo selectivo ( por ejemplo, para el IRES del VHC [ 15 – 17 ]), el progreso se ha visto limitado por la naturaleza dinámica del plegamiento del ARN y la comprensión incompleta de las estructuras de ARN y sus funciones [ 18 , 19 ]. Aquí, estudiamos la conservación de una estructura de ARN viral crítica necesaria para la replicación del ARN de ortoflavivirus y evaluamos su potencial como objetivo para la intervención antiviral.
Los genomas de los ortoflavivirus se replican en el citoplasma de las células infectadas dentro de compartimentos membranosos especializados inducidos durante la infección [ 20 , 21 ]. La síntesis de ARN viral está mediada por la actividad enzimática de la proteína no estructural NS5. Esta proteína contiene dos dominios, el dominio de ARN polimerasa dependiente de ARN (RdRp), que cataliza la síntesis de novo de ARN virales de cadena negativa y positiva [ 22 – 26 ], y el dominio de metiltransferasa (MTasa) responsable de la metilación del ARN [ 27 – 29 ]. Otras proteínas virales no estructurales también participan en el proceso de replicación del ARN [ 30 ].
Hace dos décadas, nuestro laboratorio dilucidó el mecanismo de síntesis de ARN del DENV, que requiere tres elementos de ARN esenciales dentro del genoma viral [ 31 ]. Primero, un elemento promotor conocido como tallo-bucle A (SLA), ubicado en el extremo 5′ del genoma, se une a la polimerasa viral NS5 y dirige el inicio de la síntesis de ARN [ 31 ]. Segundo, secuencias complementarias ubicadas en los extremos 5′ y 3′ del genoma median interacciones ARN-ARN de largo alcance que promueven la ciclación del genoma, un requisito previo para el inicio de la síntesis de ARN en el extremo 3′ del ARN [ 32 – 34 ]. Tercero, un tallo-bucle 3′ altamente estructurado (3′SL) sufre un reordenamiento conformacional tras la ciclación del genoma, exponiendo el extremo 3′ para servir como plantilla para el inicio de la síntesis de ARN de cadena negativa [ 35 – 39 ]. Aunque este mecanismo se definió inicialmente utilizando el modelo DENV ( Fig. 1B ), los tres elementos estructurales del ARN están presentes en todos los genomas de ortoflavivirus conocidos, lo que sugiere firmemente que el mecanismo de síntesis de ARN se conserva en todo el género.
La función de la SLA para la síntesis de ARN viral ha sido validada experimentalmente en múltiples virus, incluidos DENV, ZIKV, TBEV y WNV [ 40 – 42 ]. Varias estructuras de resolución atómica de SLA sola y en complejo con NS5 usando cristalografía de rayos X, RMN y crio-EM proporcionaron información sobre el modo de unión SLA-NS5 [ 43 – 45 ]. Las estructuras disponibles del complejo SLA-NS5 identifican dos sitios de interacción: uno en el bucle superior (TL) de la SLA que está en contacto con residuos del dominio RdRp y el otro en el extremo 5′ del ARN que interactúa con el dominio MTasa ( Fig 1C ). La unión de NS5 a la SLA conduce a la activación de la RdRp viral para la iniciación de novo de la síntesis de ARN por un mecanismo aún no claro. Dado su papel esencial en la replicación viral, examinamos la estructura y función de la SLA y su potencial como objetivo antiviral.
En este estudio, demostramos que el SLA del DENV o del ZIKV puede ser reemplazado funcionalmente por el SLA correspondiente de cualquier otro ortoflavivirus patógeno (MBFV o TBFV). Los virus quiméricos, que portan SLA heterólogos, se replican con eficiencias distintas, lo que revela requisitos tanto compartidos como específicos del virus para una replicación óptima del ARN viral. Mediante análisis detallados de estructura-función, identificamos subelementos esenciales del SLA que sustentan un modo de unión común de ARN-NS5 y proporcionamos información sobre la coevolución de subestructuras que distingue al MBFV del TBFV. También encontramos que los SLA del ISFV no son funcionales en el contexto del DENV o del ZIKV, a pesar de su capacidad para unirse a NS5. Basándonos en las características funcionales compartidas de los SLA de ortoflavivirus patógenos, examinamos una biblioteca de moléculas pequeñas centrada en el ARN para identificar compuestos que se unen al SLA del DENV. Mediante un ensayo de desplazamiento de fluoróforo, identificamos varios compuestos que presentan una inhibición significativa de la replicación del DENV a concentraciones sin citotoxicidad detectable. Se observó que uno de estos compuestos inhibe la replicación de diferentes ortoflavivirus, incluidos el ZIKV, el YFV y el virus Langat (LGTV), un TBFV. En conjunto, estos resultados identifican el SLA del ortoflavivirus como un elemento de ARN conservado y una diana prometedora para la intervención antiviral.
Resultados
Las SLA se conservan funcionalmente entre todos los ortoflavivirus patógenos.
Para evaluar las características estructurales y funcionales de las SLA de ortoflavivirus, primero realizamos alineaciones de secuencia y estructura utilizando las 5’UTR virales completas disponibles. Construimos modelos 2D y 3D de las SLA con secuencias de clados transmitidos por mosquitos, DENV y YFV (MBFV); transmitidos por garrapatas (TBFV); y subgrupos específicos de insectos, de doble huésped y clásicos (dhl-ISFV y c-ISFV, respectivamente) ( Fig. 1D ). Se observó una baja conservación general de la secuencia con pocos nucleótidos de SLA que presentaban una conservación >70% ( Fig. 1E ). A pesar de esta baja conservación de la secuencia, las alineaciones de secuencias estructurales y el modelado 3D de todas las SLA de ortoflavivirus indican un plegamiento general común. Las estructuras de las SLA incluyen un bucle de tallo superior (TSL), un tallo inferior (BS) y un bucle lateral (SL) o bucle de tallo lateral (SSL) conectados por una unión de 3 vías (3WJ) ( Fig. 1F ). Esto es consistente con la estructura previamente determinada por sonda química de ARN y determinación de estructura 3D por cristalografía de rayos X, RMN y crio-EM para DENV y ZIKV [ 43 – 49 ]. Los miembros de los c-ISFV carecen de un SSL, en su lugar poseen un SL ( Fig 1D ). Los alineamientos de secuencias estructurales indican nucleótidos invariables en regiones definidas del SLA, incluyendo el extremo 5′, el TL, ciertas posiciones del 3WJ y una protuberancia U dentro del BS ( Fig 1G ).
Motivados por la conservación estructural observada entre las SLA de diferentes ortoflavivirus, nos preguntamos si la SLA completa del DENV podría ser reemplazada funcionalmente por una SLA de virus pertenecientes al mismo grupo ecológico o a grupos distintos. Para abordar esta cuestión, utilizamos genomas reporteros de DENV y ZIKV con capacidad de replicación previamente descritos que codifican la luciferasa de Renilla (DENVRep y ZIKVRep; Fig. 2A ) [ 50 , 51 ]. La transfección de ARN virales con capuchón, transcritos in vitro, de estos constructos produce actividad de luciferasa que, medida a lo largo del tiempo, refleja la traducción del ARN de entrada y la posterior replicación del ARN. La comparación con un ARN de control defectuoso en la replicación que porta una mutación en el sitio catalítico de NS5 (NS5Mut) permite discriminar el inicio y la magnitud de la amplificación del ARN viral.
-
PNGimagen más grande
-
PELEAimagen original
A. Representación esquemática de los genomas reporteros de DENV y ZIKV (DENVRep y ZIKVRep), a la izquierda; y representación de virus quiméricos con reemplazo de la estructura SLA, a la derecha. B. Replicación de virus quiméricos en el contexto de DENVRep. Se muestra la actividad de luciferasa normalizada en función del tiempo en células BHK para ARN transfectados correspondientes a DENVRep WT, virus quiméricos y NS5Mut, como se indica en cada caso. La replicación de los virus reporteros se clasificó integrando el comportamiento temporal de la señal de luciferasa y comparaciones con controles (WT y NS5 mut) como replicativa o no replicativa. Las comparaciones estadísticas se realizaron utilizando pruebas t de Welch de dos colas, considerando tres réplicas biológicas (ver Materiales y Métodos). C. Replicación de virus quiméricos en el contexto de DENVRep en células de mosquito C6/36. Se muestra la actividad de luciferasa normalizada en función del tiempo. El análisis estadístico se realizó como en BD Replicación de virus quiméricos en el contexto de ZIKVRep. La actividad de luciferasa normalizada en función del tiempo se muestra en células BHK para los ARN transfectados correspondientes a ZIKVRep WT, virus quiméricos y NS5Mut, como se indica en cada caso. El análisis estadístico se realizó como en BE Representación esquemática del diseño de virus quiméricos en el contexto del clon infeccioso de ZIKV. F. Cuantificación de copias de ARN viral por RT-PCR a partir de partículas secretadas recolectadas en el sobrenadante de células transfectadas 3 días después de la transfección para los virus indicados en cada caso. Se muestran la media y las desviaciones estándar de dos experimentos independientes. Las copias de ARN se compararon utilizando la prueba ANOVA de una vía seguida de la prueba de comparaciones múltiples de Tukey. **** indica valor p < 0,0001; *** indica valor p ajustado < 0,001; ** indica valor p ajustado < 0,01, ns indica valor p ajustado > 0,05. G. Ensayo de inmunofluorescencia que muestra la propagación de virus ZIKV WT y quiméricos en función del tiempo en células BHK. Las células se marcaron con anticuerpos anti-NS3 específicos para ZIKV. Los símbolos inferiores representan la extensión de la propagación viral. Se indica el efecto citopático (ECP). H. Imágenes representativas de ensayos de placa formados por virus quiméricos. Para cada virus, se incluyen tres imágenes que muestran los resultados de 3 diluciones seriadas consecutivas del sobrenadante obtenido a las 72 horas posteriores a la transfección. Imagen de mosquito de clip art de dominio público tomada de NIH Bio Art. https://bioart.niaid.nih.gov/bioart/162 .
Para generar construcciones quiméricas de DENVRep, el SLA completo de DENV fue reemplazado con SLA derivados de MBFV emergentes y reemergentes que representan riesgos significativos para la salud humana, incluidos ZIKV, JEV, virus Aroa (AROV), virus de la encefalitis de San Luis (SLEV), virus Usutu (USUV), virus del Nilo Occidental (WNV), virus Ilhéus (ILHV), virus Rocio (ROCV) y virus de la fiebre amarilla (YFV). También se emplearon SLA de ortoflavivirus humanos patógenos transmitidos por garrapatas, incluidos el virus de la encefalitis transmitida por garrapatas (TBEV) y el virus Powassan (POWV). Además, para tener en cuenta la diversidad ecológica, incluimos SLA del virus Nhumirin (NHUV), un dhl-ISFV, y del flavivirus Aedes (AeFV), un c-ISFV ( Fig. 2B ).
Se tuvieron en cuenta varias consideraciones durante la construcción de quimeras. Primero, se prestó especial atención a la unión de secuencia entre el SLA heterólogo y el genoma del DENV para asegurar que el dinucleótido AG conservado en el extremo 5′ del genoma no participara en el apareamiento de bases dentro del tallo inferior del SLA [ 52 ]. Además, estudios previos han demostrado que la actividad MTasa (metilación N-7) de NS5 de ciertos MBFV requiere nucleótidos específicos en las posiciones dos y tres del genoma viral [ 53 ]. El análisis de secuencia reveló variabilidad en la tercera posición de nucleótido: mientras que DENV, ZIKV, AROV, WNV y YFV conservan una U, SLEV, JEV, USUV, ROCV e ILHV contienen una A en esta posición. Para asegurar la compatibilidad entre construcciones quiméricas, se mantuvo una U en la tercera posición de nucleótido. Para obtener más detalles sobre la construcción de virus quiméricos, consulte la información complementaria ( Fig. S1 ).
Los ARN virales con capuchón correspondientes a DENVRep WT, el control NS5Mut defectuoso en la replicación y cada uno de los constructos quiméricos se generaron mediante transcripción in vitro utilizando un promotor T7 modificado para asegurar la secuencia viral auténtica del extremo 5′ GpppAGU, se cuantificaron y se transfectaron en células BHK. La actividad de la luciferasa se midió a las 4, 24, 48 y 72 hpt. Todas las quimeras que contenían SLA derivados de ortoflavivirus patógenos, incluidos YFV y los TBFV, mostraron evidencia clara de replicación del ARN viral cuando se compararon con el control NS5Mut ( Fig. 2B ). La actividad de la luciferasa normalizada a las 72 hpt fue significativamente mayor que la del control NS5Mut. Varios virus quiméricos exhibieron un fenotipo replicativo pero subóptimo, como las quimeras que contenían SLA de ILHV, ROCV, JEV y YFV ( Fig. 2B ). Para el análisis estadístico, consulte Material y Métodos. Por el contrario, las quimeras que portaban SLA del ISFV clásico y de tipo huésped dual (NHUV y AeFV, respectivamente) no lograron replicarse, mostrando niveles de luciferasa que no eran significativamente diferentes a los del control NS5Mut ( Fig. 2B ).
Dado que el ISFV se replica exclusivamente en células de insectos, examinamos si la falta de replicación observada para las quimeras NHUV y AeFV se debía a la restricción de la célula huésped. Para ello, se transfectaron ARN virales con capuchón de DENVRep WT, NS5Mut, la quimera ZIKV SLA y las quimeras ISFV (NHUV y AeFV) en células de mosquito C6/36. Las mediciones de luciferasa revelaron una replicación eficiente de la quimera ZIKV SLA, mientras que ambos ISFV permanecieron incapaces de replicarse ( Fig. 2C ). Estos resultados indican que la incapacidad de las SLA de ISFV para sustentar la replicación en el contexto del DENV no se debe a la especificidad de la célula huésped.
Nuestros datos sugieren que el SLA de cualquier ortoflavivirus patógeno puede sustentar la replicación del ARN viral en el contexto del DENV, lo que es consistente con la conservación funcional de esta estructura de ARN. Para validar aún más esta conclusión, generamos quimeras análogas en el contexto de ZIKVRep. Se utilizaron SLA de ortoflavivirus representativos para reemplazar el SLA nativo de ZIKV, incluyendo SLA de DENV, AROV, YFV, TBEV, POWV, NHUV y AeFV. De acuerdo con los resultados obtenidos en el contexto del DENV, las quimeras con SLA de MBFV y TBFV sustentaron la amplificación del ARN en el contexto del ZIKV, mientras que las quimeras con SLA de ISFV fueron inactivas ( Fig. 2D ).
Para confirmar aún más nuestros hallazgos obtenidos con los sistemas reporteros DENV y ZIKV, analizamos la función de SLA en el contexto de virus infecciosos. Se generaron virus quiméricos utilizando un clon infeccioso de ZIKV previamente construido en nuestro laboratorio [ 50 ], en el que el SLA nativo de ZIKV fue reemplazado por el SLA de DENV, YFV, TBEV, NHUV o AeFV ( Fig. 2E ). Los ARN virales con capuchón se transfectaron en células, y la replicación y propagación viral se evaluaron mediante la cuantificación por RT-PCR del ARN en partículas virales secretadas, mediante ensayos de inmunofluorescencia (IFA) que detectan la proteína viral NS3, y mediante ensayos de placa ( Fig. 2F , 2G y 2H ). Los virus que contenían SLA de DENV, YFV o TBEV fueron capaces de replicarse y propagarse, aunque con una eficiencia reducida en comparación con el ZIKV WT ( Fig. 2F ). Por el contrario, las quimeras que portaban SLA de ISFV no lograron replicarse, sin mostrar proteínas virales detectables ( Fig. 2G ).
En conjunto, estos resultados demuestran que las SLA derivadas de los virus patógenos MBFV o TBFV conservan su función y permiten la replicación del ARN viral en contextos genómicos heterólogos. A pesar de la divergencia de secuencias, estas SLA mantienen características estructurales y funcionales compartidas que posibilitan una interacción productiva con la maquinaria de replicación viral del DENV y el ZIKV, lo que concuerda con un mecanismo conservado de reconocimiento de SLA por la proteína NS5 durante la metilación de la caperuza y la síntesis del ARN viral.
Relaciones estructura-función entre las SLA de ortoflavivirus
Descubrimos que las SLA de MBFV y TBFV permiten la replicación del ARN viral en el contexto del DENV y el ZIKV, aunque con diferentes eficiencias de replicación, mientras que las de ISFV no son compatibles con el DENV ni el ZIKV. Para comprender mejor las relaciones estructura-función de los subelementos de las SLA, analizamos características específicas de grupo y de virus en los modelos 2D y 3D de las SLA de diferentes ortoflavivirus.
Primero examinamos las propiedades de la TL. Se ha demostrado que la secuencia de nucleótidos en la TL es crítica para la actividad de la polimerasa NS5 dependiente de DENV SLA, demostrada por la síntesis de ARN de novo in vitro e infecciones virales [ 31 , 46 ]. Los estudios funcionales también asignaron un papel esencial a las secuencias TL en WNV [ 54 ]. Los análisis estructurales de la DENV SLA en complejo con NS5 identificaron contacto directo entre el dominio del pulgar de la RdRP y nucleótidos específicos de la TL ( Fig 1C ) [ 43 , 45 , 55 ]. Por lo tanto, examinamos con más detalle la conservación de la secuencia TL entre las SLA de ortoflavivirus que representan los diferentes grupos ecológicos. Se observó una secuencia consenso YVGN, en la que Y representa una pirimidina (C/U), V cualquier nucleótido excepto U, y N cualquier nucleótido ( Fig 3A ). Los TL se restringieron a ocho secuencias únicas en aislados naturales (solo el 3% del espacio de secuencias posible): CAGU, CAGA, CAGG, CAGC, UAGU, CGGA, UGGU, CCGU ( Fig. 3A ). CAGN fue el motivo más prevalente (59% de las secuencias), seguido de UAGU ( Fig. 3A ). Curiosamente, la variación de la secuencia TL no se correlacionó con la clasificación del grupo ecológico. Por ejemplo, UAGU se encontró en un MBFV (WNV), un TBFV (POWV) y un ISFV (NHUV) ( Fig. 3A ).
-
PNGimagen más grande
-
PELEAimagen original
A. Conservación de la secuencia de TL de MBFV, TBFV e ISFV. Logotipo obtenido a partir de la alineación que muestra la frecuencia de nucleótidos individuales. A la derecha, tabla que muestra la secuencia de nucleótidos TL para cada virus. Se indican los miembros de los grupos MBFV, TBFV e ISFV (clásicos y de tipo huésped dual). B. Impacto de las secuencias TL en la replicación del ARN de ZIKVRep. Se muestra la actividad de luciferasa normalizada en función del tiempo para el WT, el mutante ZIKVRep y el NS5Mut en células BHK. La secuencia TL para cada mutante se indica en la parte superior. El código de color corresponde a cada secuencia TL como se indica en la tabla anterior. La replicación de los virus reporteros se clasificó integrando el comportamiento temporal de la señal de luciferasa y comparaciones con controles (WT y NS5 mut) como replicativa o no replicativa. Las comparaciones estadísticas se realizaron utilizando pruebas t de Welch de dos colas, considerando tres réplicas biológicas (ver Materiales y Métodos). C. Instantánea de la simulación MD que muestra los contactos entre los nucleótidos TL y los residuos del dominio del pulgar RdRp de NS5 conservados durante las trayectorias MD de 1 microsegundo (3 réplicas). Se indican los nucleótidos SLA TL C30, A31, G32. Se muestra la región del pulgar. Los residuos con 100% de conservación entre los ortoflavivirus están marcados con un asterisco. D. Instantáneas de simulaciones MD superpuestas para mostrar las conformaciones de las nucleobases SLA TL (C30, A31, G32 y U33). La interacción entre la primera y la tercera base (línea discontinua) se mantiene durante las tres simulaciones de microsegundos. E. Longitud de TS, BT y SS agrupadas según el grupo ecológico. Las longitudes se compararon utilizando la prueba no pareada de Mann-Whitney. **** indica un valor p ajustado <0,0001; ns indica un valor p ajustado >0,05. F. Impacto de diferentes SSL en la replicación viral. Actividad de luciferasa normalizada en función del tiempo para ZIKVRep WT, quimeras y NS5Mut en células BHK como se indica en cada caso. Representación de SLA con el SSL correspondiente en diferentes colores. El análisis estadístico se realizó como en BG Instantáneas superpuestas de la simulación MD que resaltan la posición del apilamiento SS y TS-SS observado en el 97% de las configuraciones. H. Fracción de secuencias SLA con al menos 1 modelo 3D con apilamiento coaxial TS-SS en cada grupo ecológico. En la parte inferior una representación esquemática del 3WJ y el apilamiento coaxial TS-SS.
Para investigar el requerimiento funcional de las secuencias TL, utilizamos ZIKVRep. Diseñamos ARN virales quiméricos en los que la secuencia TL de tipo silvestre (WT) fue reemplazada por la de virus representativos: DENV, YFV, TBEV, NHUV o por una secuencia UCAG no consensuada (NC) que carece de la G conservada en la tercera posición, la cual no se observa en la naturaleza ( Fig. 3B ). La secuencia TL de AeFV es la misma que la de NHUV (UAGU) ( Fig. 3A ). Los ARN virales cuantificados de ZIKVRep WT y los virus quiméricos se transfectaron en células BHK junto con el control NS5Mut. Los ARN con SLA que albergaban TL encontrados en aislados naturales permitieron una replicación eficiente del ARN viral, como lo indica el aumento de la actividad de la luciferasa con el tiempo. Por el contrario, el ARN viral que contenía la TL NC no se replicó, mostrando una cinética de luciferasa comparable a la del control NS5Mut ( Fig. 3B ). Estos hallazgos indican que la TL del SLA tolera una variabilidad de secuencia limitada, con solo un conjunto restringido de motivos naturales que respaldan la replicación del ARN viral, lo cual es independiente de los grupos ecológicos virales.
La G invariable encontrada en la tercera posición en la TL de todos los ortoflavivirus coincide con interacciones específicas entre esta nucleobase con R770 y K841 en la región del pulgar de RdRp [ 43 , 45 ]. Para explorar la estabilidad de estas interacciones específicas, realizamos dinámica molecular (MD) del complejo NS5-SLA. Se encontró que la interacción específica nucleobase-cadena lateral del tercer nucleótido TL (G32) con R770 era estable a lo largo de las tres simulaciones de microsegundos, con una distancia entre el oxígeno carbonilo de la base guanina y el carbono central del grupo guanidinio de la cadena lateral de 3,6 +/- 0,6 Å ( Figs 3C y S2 ). Por el contrario, la interacción entre la nucleobase y K841 no se conservó en ninguna de las tres simulaciones (distancia promedio nucleobase-cadena lateral de 7 +/- 1 Å) ( Fig S2 ). Otros residuos dentro del dominio del pulgar de la RdRp que establecen interacciones estables con el esqueleto de ARN de los primeros tres nucleótidos de la TL se destacan en la Fig. 3C . Desde la perspectiva de NS5, se observó conservación de residuos entre todos los ortoflavivirus. Las cadenas laterales de aminoácidos de la RdRp que mantienen interacciones estables con la TL (R856, R770, Y838 y R888, Fig. 3C ) están 100% conservadas entre los ortoflavivirus. La conservación parcial de la secuencia y la función de los primeros y segundos nucleótidos de la TL podría explicarse por su participación en interacciones proteína-ARN mediadas a través del esqueleto de ARN en lugar de las nucleobases. La conformación del esqueleto de ARN se estabiliza por una interacción entre la primera y la tercera nucleobase que persiste durante las tres trayectorias de MD de microsegundos ( Fig. 3D ). Esto es consistente con la variabilidad de secuencia restringida de la primera posición a C o U; Ambas nucleobases poseen un grupo carbonilo en la posición C2 de su anillo de pirimidina, el cual participa en un enlace de hidrógeno con la guanina conservada en la tercera posición. Por otro lado, el cuarto nucleótido adopta múltiples conformaciones que se alejan de la superficie de la proteína ( Fig. 3D ), lo cual concuerda con la gran variabilidad natural observada en esta posición. En conjunto, los perfiles de replicación de los genomas de ZIKVRep que portan diferentes secuencias TL naturales, junto con los análisis estructurales y de secuencia, respaldan un modo de interacción conservado entre la TL y el dominio del pulgar de la RdRP en los ortoflavivirus.
Luego, investigamos las estructuras y funciones de los tallos que forman el SLA. Dentro del BS, se identificó originalmente una protuberancia de U en el DENV como esencial para la replicación del ARN. La sustitución U/A en esta posición resultó en virus revertantes que restauraron la secuencia [ 46 ]. Observamos una conservación absoluta de la protuberancia de U en diferentes virus, lo que respalda una función importante ( Fig 1D ). Una diferencia importante entre los SLA de ortoflavivirus es la presencia y longitud del bucle lateral del tallo (SSL) ( Fig 1D ). Los modelos 2D predichos de los SLA de virus patógenos muestran un SSL, mientras que los SLA de c-ISFV muestran un bucle lateral (SL), lo que representa una diferencia importante entre distintos grupos ecológicos virales. El tallo lateral (SS) podría estar en un equilibrio dinámico con un SL basado en observaciones previas, incluida la reactividad intermedia en ensayos SHAPE [ 43 , 45 – 49 ]. En los MBFV, la longitud del SS varía de 3 a 5 pares de bases, mientras que en los TBFV, puede extenderse hasta 21 pares de bases ( Fig. 3E ). Para evaluar la relevancia funcional de la presencia y longitud de un SS, generamos ARN quiméricos ZIKVRep reemplazando el SSL del ZIKV con el de dos TBFV, POW u OHFV, así como YFV, y una secuencia SL de c-ISFV ( Fig. 3F ). Los ARN virales se cuantificaron y se transfectaron en células BHK junto con el control no replicativo. La actividad de la luciferasa medida en función del tiempo indicó que los constructos con SLA que contenían cualquier SSL apoyaban la amplificación del ARN viral como el virus WT. Por el contrario, el ARN viral con el SLA que portaba la secuencia SL de c-ISFV no mostró signos de replicación del ARN ( Fig. 3F ). El análisis estructural del complejo SLA con NS5 indica que el SSL sobresale de la superficie de la proteína, lo que explica por qué la longitud de este elemento no afecta la función del SLA en la amplificación del ARN viral ( Fig. 3G ). Sin embargo, la presencia de este elemento parece ser crucial.
Se observó que el apilamiento coaxial del SS con el TS en la estructura DENV SLA-NS5 era estable en las tres simulaciones de dinámica molecular de microsegundos independientes (97 % de las conformaciones). Curiosamente, este apilamiento coaxial del TS y el SS también se observó en los modelos 3D de SLA de MBFV y TBFV, con la excepción de YFV ( Fig. 3H ). Estos resultados sugieren que el apilamiento SS-TS es una característica común que podría ser importante para la función de SLA.
Los tres tallos convergen en el 3WJ, lo cual es importante para definir la disposición arquitectónica 3D del SLA. En el 3WJ, cortos enlazadores monocatenarios conectan las tres hélices (uniones J 1,2 , J 2,3 y J 1,3 , Fig 4A ) y las longitudes de los enlazadores influyen fuertemente en la geometría de la unión, el apilamiento coaxial y la flexibilidad del tallo. Los 3WJ se pueden clasificar como Tipos A, B o C según qué hélices se apilan coaxialmente y según las longitudes de estas uniones [ 56 ]. El 3WJ del SLA de DENV2 pertenece al Tipo C, en el que el TS y el SS están apilados, el enlazador inferior-superior J 1,2 es más largo que el enlazador lateral-inferior J 1,3 , y el enlazador superior-lateral J 2,3 está ausente (0 nt). La alineación de secuencias de estructura secundaria conservada en los SLA de MBFV y TBFV reveló motivos 3WJ específicos de grupo, con YFV mostrando un patrón único ( Fig. 4B ). Los SLA de MBFV muestran que J 1,2 es GAG, GAA o GGA, J 2,3 está ausente y J 1,3 es una sola A conservada. En el caso de YFV, J 1,2 es CUAA, J 2,3 está ausente y J 1,3 es AAA. En TBFV, J 1,2 sigue una secuencia consenso GHR (donde H es cualquier cosa excepto G, y R una purina), J 2,3 está ausente y J 1,3 contiene dos nucleótidos, GA o UA. El 3WJ de dhl-ISFV muestra un patrón similar al de MBFV, pero J 1,2 es GCA, GCG o GGA ( Fig. 4B ).
-
PNGimagen más grande
-
PELEAimagen original
A. Representación esquemática del 3WJ, que indica las tres hélices y los conectores monocatenarios que las unen: J 1,2 ; J 2,3 y J 1,3 . B. Secuencias del 3WJ en cada miembro de los diferentes grupos como se indica, MBFV, TBFV y dhl-ISFV. A la derecha, la secuencia consenso del 3WJ y una representación de la estructura para cada grupo, como se indica. C. Actividad de luciferasa normalizada en función del tiempo para ZIKVRep WT, quimeras 3WJ y NS5mut en células BHK como se indica. La replicación de los virus reporteros se clasificó integrando el comportamiento temporal de la señal de luciferasa y las comparaciones con muestras de control (WT y NS5 mut) se clasificaron como replicativas o no replicativas. Las comparaciones estadísticas se realizaron utilizando pruebas t de Welch de dos colas, considerando tres réplicas biológicas (ver Materiales y Métodos). D. Ensayo de inmunofluorescencia que muestra la propagación en función del tiempo del ZIKV WT y los virus que portan el 3WJ del YFV y el TBEV como se indica, las células BHK se marcaron con anticuerpos anti-NS3 del ZIKV. Se indica el efecto citopático (ECP). E. Ángulos de pseudotorsión η y θ correspondientes a los nucleótidos 3WJ de J 1,2 de los modelos 3D SLA y del DENV2 durante la simulación MD. F. Actividad de luciferasa normalizada en función del tiempo para ZIKVRep WT, NS5Mut y virus quiméricos que portan el 3WJ o 3WJ + SS del TBEV como se indica. La comparación entre el TBEV 3WJ y el TBEV 3WJ + SS a las 48 y 72 hpt se realizó utilizando pruebas t de Welch de dos colas. *** indica un valor p ajustado <0,001. G. Representación que indica la distancia entre el TP y el extremo 5′ del ARN en el complejo RNP. H. Distribución de la distancia 5′-TL (en Å) en modelos 3D de SLA de los grupos ecológicos MBFV (azul), TBFV (naranja), dh-ISFV (rosa oscuro) y c-ISFV (rosa claro). Cada histograma se calculó utilizando la función de gráfico de histograma de seaborn con 40 bins y se normalizó de manera que el área total sea 1. Se incluye un gráfico de densidad de kernel para facilitar la visualización de las distribuciones (ancho de banda determinado utilizando la regla de Scott) [ 58 ]. La región del gráfico de densidad de kernel del grupo MBFV sombreada en azul oscuro resalta los valores para YFV (63-73 Å). Las líneas verticales hacen referencia a las distancias 5′-TL correspondientes de la estructura NS5-SLA (PDB 8gzp, 59 Å, línea negra continua) [ 45 ], y de las simulaciones de dinámica molecular del complejo (52 ± 2 Å, línea discontinua vertical y región sombreada circundante).
Para evaluar el papel del 3WJ dentro de la estructura SLA en la replicación viral, generamos un nuevo ZIKVRep quimérico reemplazando solo la secuencia 3WJ del SLA del ZIKV con la del YFV (3WJ-YFV ZIKVrep) o TBEV (3WJ-TBEV ZIKVrep). Las predicciones de la estructura secundaria de estos SLA quiméricos sugirieron que el plegamiento general del SLA se conservó. El ARN del ZIKVRep WT, las dos quimeras y el NS5Mut ZIKVrep, se cuantificaron y transfectaron en células BHK. Los niveles de expresión de luciferasa mostraron que el virus que portaba el 3WJ-TBEV retuvo una capacidad de replicación parcial, mostrando una reducción de ~ 2 log en relación con el ZIKVRep WT a las 24 horas, mientras que el ARN con el 3WJ-YFV mostró una replicación mínima ( Fig. 4C ). Los resultados sugieren que la secuencia y estructura 3WJ es un elemento sensible para la función SLA. Para confirmar esta observación, utilizamos las mismas mutaciones 3WJ pero en el contexto del clon infeccioso ZIKV y evaluamos si los virus quiméricos (3WJ-YFV ZIKV y 3WJ-TBEV ZIKV) eran capaces de replicarse y propagarse en células cultivadas. Seguimos la propagación viral mediante IFA contra la proteína viral NS3. Mientras que el ZIKV WT mostró una monocapa completamente infectada a las 48 hpt y a las 96 hpt la mayor parte de la monocapa mostró efecto citopático (ECP), se observaron signos de propagación de ambas quimeras a las 96 hpt, lo que sugiere una replicación ineficiente de estos virus, en concordancia con los datos obtenidos con el ZIKVRep ( Fig. 4D ).
Para caracterizar estructuralmente el 3WJ, analizamos los ángulos de pseudotorsión η (eta) y θ (theta) en los modelos 3D de los SLA correspondientes a los nucleótidos de J 1,2 . Los ángulos η y θ representan la geometría del esqueleto en dos dimensiones y permiten la identificación de motivos estructurales [ 57 ]. Observamos que los nucleótidos de J 1,2 generalmente caen en las mismas regiones del espacio η-θ dentro de cada grupo ecológico, lo que indica una conformación similar de los 3WJ ( Fig. 4E ). Para el grupo MBFV, este patrón coloca el primer, segundo y tercer nucleótido de J 1,2 en las regiones helicoidal, de giro apilado y de torsión de base, respectivamente. Esta conformación coincide con la del DENV4 SLA en la estructura unida a NS5 [ 45 ], y cada uno de los nucleótidos permaneció en su región correspondiente durante las tres simulaciones de MD de microsegundos. El dh-ISFV 3WJ también coincide en estas regiones del gráfico η-θ, mientras que el YFV se agrupó por separado. El MBFV y el TBFV ocupan regiones distintas en el espacio η–θ, lo que respalda la idea de que la arquitectura 3WJ divergió entre estos grupos ecológicos ( Fig. 4E ).
Observamos que la introducción del SLA del TBEV en el contexto del ZIKV retrasa la replicación viral significativamente menos que la sustitución únicamente de los nucleótidos 3WJ (compárense las figuras 2D y 4C ). Varias explicaciones podrían justificar esta diferencia. Una posibilidad es que elementos estructurales adicionales dentro del SLA del TBEV compensen el impacto del 3WJ heterólogo en el virus quimérico. Dado que el SSL difiere notablemente entre los MBFV y los TBFV, planteamos la hipótesis de que estos dos subelementos del SLA (el SSL y el 3WJ) podrían haber coevolucionado dentro de distintos grupos ecológicos.
Para probar esta hipótesis, generamos un nuevo ZIKVRep quimérico en el que tanto el 3WJ como el SSL del ZIKV fueron reemplazados por los elementos correspondientes del TBEV. Se generaron ARN virales con capuchón y se transfectaron en células, y se monitorizó la replicación mediante la actividad de la luciferasa. La quimera que contenía ambos elementos del TBEV se replicó de forma significativamente más eficiente, aproximadamente dos órdenes de magnitud superior, que el virus que portaba solo el 3WJ del TBEV ( Fig. 4F ). Estos resultados indican que, si bien las secuencias 3WJ de los SLA han divergido entre virus de diferentes grupos ecológicos, la coevolución con otros subelementos SLA puede preservar la función del promotor.
La estructura NS5–SLA disponible indica que el dominio MTasa contacta el extremo 5′ (BS), mientras que la RdRp se une a la TL. Por lo tanto, la orientación relativa de TSL y BS, y la distancia entre el extremo 5′ y la TL, son probablemente determinantes críticos de la unión de NS5 ( Fig. 4G ). En la estructura de crio-EM del complejo DENV NS5–SLA, esta distancia es aproximadamente 59 Å y converge a un valor promedio de 52 + /- 2 Å durante las simulaciones de MD. Las distribuciones de esta distancia en modelos 3D de SLA libres de ortoflavivirus patógenos (MBFV y TBFV) están centradas cerca de estos valores, con valores promedio de 53 y 54 Å para los grupos MBFV y TBFV, respectivamente, excepto para YFV con una distancia predicha centrada en 65 Å ( Fig. 4H ). Por el contrario, las SLA del grupo c-ISFV muestran una distribución marcadamente distinta centrada en ~91 Å, sin solapamiento con los grupos patógenos ( Fig. 4H ). En conjunto, estos resultados indican que la distancia 5′-TL de las SLA es otra característica estructural común entre los ortoflavivirus patógenos.
En conjunto, nuestros análisis funcionales y estructurales indican que la replicación viral productiva requiere no solo estructuras conservadas y secuencias específicas dentro de los subelementos SLA individuales, sino también su organización coordinada. Si bien ciertas características de SLA han divergido de manera específica para cada grupo, la combinación de estos elementos mantiene una arquitectura global conservada de SLA que permite el reconocimiento heterólogo de NS5 y la replicación del ARN viral.
La unión de las SLA del ISFV a NS5 no es suficiente para sustentar la replicación del ARN.
Los miembros del grupo ISFV se dividen en dos subgrupos filogenéticamente distintos. Mientras que el subgrupo con doble huésped está genéticamente más emparentado con los MBFV, el subgrupo clásico está más distantemente relacionado, una divergencia que se refleja en la estructura de sus SLA ( Fig. 1A y 1D ). Los análisis comparativos revelaron características estructurales y funcionales distintas entre los subelementos SLA de estos virus, lo que podría explicar la falta de funcionalidad de SLA observada en el contexto del DENV y el ZIKV.
Para investigar más a fondo la base de la ausencia de replicación observada en los MBFV quiméricos que portan SLA derivados de ISFV, examinamos la capacidad de estos SLA para formar complejos heterólogos con DENV NS5. Las interacciones proteína-ARN se analizaron utilizando ensayos de desplazamiento de movilidad electroforética (EMSA). Se produjo DENV NS5 recombinante purificado y se generaron moléculas de ARN correspondientes a los SLA de DENV, YFV, TBEV, NHUV y AeFV. Los ARN SLA se utilizaron a una concentración fija (10 nM), mientras que NS5 se tituló de 10 a 120 nM ( Fig. 5A ).
-
PNGimagen más grande
-
PELEAimagen original
A. Interacción SLA-NS5 por EMSA. Geles representativos que muestran las propiedades de unión de la proteína NS5 purificada a SLA de DENV, TBEV, YFV, AeFV, NHUV y un ARN no relacionado como se indica. Se muestra la movilidad del SLA libre y el complejo RNP. La concentración de NS5 se indica en la parte superior, y la concentración de SLA es de 10 nM. Un gráfico con curvas de unión de NS5 a los SLA indicados basado en la fracción de ARN unido del EMSA se muestra en la parte inferior derecha. Se ajustó un modelo de curva logística sigmoidal de cuatro parámetros a los datos. B. Búsqueda de mutaciones adaptativas de virus quiméricos no replicantes que portan SLA de ISFV. En la parte superior, representación esquemática del procedimiento experimental que muestra pases seriados (P1, P2, P3 y P4). En la parte inferior, imágenes de ensayos de inmunofluorescencia de células de mamíferos transfectadas con ARN infeccioso de ZIKV y los virus quiméricos (NHUV ZIKV y AeFV ZIKV) en función del tiempo. NHUV ZIKV se replica después de pases seriados. La secuencia del virus rescatado revela una mutación en el tallo superior, A26U. A la derecha, la estructura secundaria de NHUV SLA indica la ubicación de la mutación A26U. C. La mutación obtenida es suficiente para sustentar la replicación del ARN viral (NHUV-R ZIKV). Se muestran los niveles normalizados de luciferasa en función del tiempo para ZIKVRep WT, NHUV-ZIKVRep, las quimeras NHUV-R ZIKVRep y NS5Mut. La comparación entre el virus quimérico de NHUV y NHUV-R a las 48 y 72 hpt se realizó utilizando pruebas t de Welch de dos colas. ** indica un valor p ajustado <0,01; * indica un valor p ajustado <0,05.
Como control, el EMSA realizado con el DENV SLA reveló la formación de un complejo de ribonucleoproteína (RNP) definido, consistente con informes previos [ 31 , 59 ]. En la mayoría de las concentraciones de NS5, se observó un único complejo discreto, mientras que en concentraciones más altas de NS5 aparecieron complejos de migración más lenta, consistente con la unión de múltiples moléculas de NS5 [ 43 , 54 , 59 ]. Se detectaron complejos RNP comparables con SLA de DENV, TBEV, YFV y NHUV. Por el contrario, aunque el AeFV SLA también fue capaz de formar un complejo con DENV NS5, su afinidad de unión aparente se redujo notablemente ( Fig 5A ). Se incluyó un ARN no relacionado de longitud idéntica como control para interacciones no específicas; en este caso, se detectó la unión de NS5, como se esperaba dada su naturaleza de unión al ARN, pero no se formó ningún complejo RNP definido.
En conjunto, estos resultados sugieren que la proteína SLA del NHUV se une a la proteína NS5 del DENV con una afinidad aparente comparable a la de la proteína SLA del DENV, mientras que la proteína SLA del AeFV muestra una unión sustancialmente más débil. Estos datos sugieren que una interacción de alta afinidad entre la proteína SLA del NHUV y la proteína NS5 no es suficiente para sustentar la replicación del ARN en el contexto de virus quiméricos.
Para investigar más a fondo la incapacidad de los SLA del ISFV para funcionar como promotores de la replicación del ARN del MBFV, buscamos mutaciones adaptativas que surgieran durante el paso seriado en cultivo celular. Las células se transfectaron con ARN infecciosos del ZIKV que portaban los SLA del NHUV o del AeFV, junto con el ZIKV de tipo salvaje como referencia, y se sometieron a pasos sucesivos cada 72 horas durante un total de 15 días. La replicación viral se monitorizó mediante IFA ( Fig. 5B ). La quimera que portaba el SLA del NHUV mostró signos de propagación viral en el paso 4 (12 días), mientras que no se detectó propagación para el virus que portaba el SLA del AeFV, incluso cuando los experimentos se extendieron a 20 días.
Para determinar si la replicación de la quimera NHUV SLA estaba asociada con mutaciones adaptativas, se extrajo ARN viral de los sobrenadantes recolectados en el paso 4, se amplificó mediante RT-PCR y se secuenciaron las regiones codificantes 5′UTR y NS5. Experimentos independientes identificaron una sustitución de un solo nucleótido dentro de NHUV SLA. Esta mutación se mapeó en el TS de la estructura SLA, que podría formar un par de bases adicional ( Fig. 5B ). Para probar directamente si esta mutación restauraba la función de SLA en el contexto del ZIKV, el cambio identificado se introdujo en el constructo no replicativo ZIKVRep-NHUV ( Fig. 5C ). Los ARN con capuchón correspondientes a WT ZIKVRep, la quimera NHUV SLA original y NHUV SLA rescatado (NHUV-R) se transfectaron en células, y la replicación del ARN se monitorizó mediante la actividad de la luciferasa. Los datos muestran que el constructo NHUV-R apoyó la replicación del ARN viral, mostrando niveles de luciferasa cerca de 3 órdenes de magnitud más altos que la quimera parental a las 72 h ( Fig. 5C ).
En conjunto, estos resultados demuestran que la estructura SLA del NHUV puede adquirir compatibilidad funcional con el complejo de replicación del MBFV mediante una adaptación estructural mínima, mientras que la SLA del AeFV carece de características esenciales que no se rescataron mediante la evolución impulsada por el cultivo celular.
Pequeñas moléculas que se unen a SLA como inhibidores de la replicación viral
Para identificar ligandos de moléculas pequeñas de SLA, examinamos una biblioteca dirigida a ARN de moléculas pequeñas comerciales (DRTL) y moléculas pequeñas sintéticas reportadas en trabajos recientes [ 60 – 62 ]. Se determinó que un ensayo de desplazamiento de indicador (IDA) utilizando el colorante TOPRO-1 era adecuado para el cribado basado en una puntuación Z’ de 0,54 (una puntuación Z’ > 0,4 es aceptable) ( Fig. 6A ). Las placas que contenían el colorante TOPRO-1 y DENV2 SLA se trataron con 10 µM de molécula pequeña y se calculó el porcentaje de desplazamiento del colorante. Los datos de tres cribados independientes se promediaron y los resultados positivos se identificaron como moléculas pequeñas con valores de porcentaje de desplazamiento del indicador fluorescente (%FID) mayores del 20% (46 resultados positivos, tasa de resultados positivos del 4,7%). De estos resultados positivos, se seleccionaron 10 ligandos para pruebas adicionales basadas en %FID y frecuencia de resultados positivos en otros ensayos.
-
PNGimagen más grande
-
PELEAimagen original
A. Izquierda: Representación del ensayo de desplazamiento del indicador utilizando el colorante TOPRO-1. Cuando el indicador (estrella) se une al ARN, la señal se potencia. El desplazamiento del indicador por una molécula pequeña competitiva (esfera) disminuye la señal. El ensayo se utilizó para examinar la biblioteca dirigida al ARN contra DENV2 SLA. Derecha: Gráfico de cascada de los resultados del cribado a 10 µM de molécula pequeña. B. Se utilizaron SM seleccionados que se unen a DENV2 SLA para evaluar el impacto en la replicación de DENVRep. En la parte superior, representación esquemática de la producción de partículas virales de DENVRep y cribado de la replicación del ARN viral midiendo la actividad de la luciferasa. En la parte inferior, se muestran las curvas de inhibición viral y viabilidad celular para cada molécula, como se indica. C. Inhibición viral y selectividad de los SM seleccionados. Se muestran gráficos que representan la IC 50 aparente y la CC 50 /IC 50. D. Inhibición de DENV2 WT por DMA-176 y DMA-177. En la parte superior, representación esquemática del ensayo y, en la parte inferior, gráficos que muestran la reducción del título viral (verde) y la viabilidad celular (gris) en función de la concentración de SM. Las curvas son la media de tres experimentos independientes. Derecha: Estructuras de DMA-176 y DMA-177. E. Efecto de DMA-176 y DMA-177 en la infección de ZIKV WT, YFV 17D y LGTV. Los títulos virales se midieron en función de la concentración de SM. También se muestra la viabilidad celular (gris). Se ajustaron modelos de curva logística sigmoidal de cuatro parámetros a los datos de luciferasa, títulos virales y viabilidad celular para determinar la IC 50 y CC 50 aparentes . Los dibujos utilizados en esta figura incluyen imágenes prediseñadas de dominio público tomadas de NIH Bio Art: https://bioart.niaid.nih.gov/bioart/7 y https://bioart.niaid.nih.gov/bioart/156 .
Los ligandos seleccionados se evaluaron posteriormente para determinar su actividad antiviral. La evaluación inicial se realizó utilizando el sistema DENVRep, que produce partículas infecciosas que codifican luciferasa. Se generaron cepas virales en células de mosquito y se utilizaron para infectar células humanas en presencia de concentraciones crecientes de SM ( Fig. 6B ). La replicación viral se cuantificó mediante la actividad de la luciferasa, y la citotoxicidad se evaluó en paralelo para cada compuesto. Se determinaron las concentraciones inhibitorias medias aparentes (aIC₅₀) y las concentraciones citotóxicas aparentes (aCC₅₀) ( Fig. 6C ). Las moléculas que mostraron los índices de selectividad más altos se seleccionaron para un análisis posterior (dimetil amilorida DMA-176 y DMA-177) utilizando DENV WT.
Las células humanas A549 se infectaron con DENV a una multiplicidad de infección (MOI) de 0,1 en presencia de concentraciones crecientes de DMA-176 o DMA-177. La producción de partículas virales en los sobrenadantes de cultivo se cuantificó mediante ensayo de placa, y la citotoxicidad de los compuestos se evaluó utilizando CellTiter-Glo. Ambos compuestos inhibieron la replicación del DENV a concentraciones submicromolares, con valores de aIC₅₀ de 0,3 ± 0,2 μM para DMA-176 y 0,8 ± 0,1 μM para DMA-177.
A continuación, examinamos si estos compuestos inhibían la replicación de otros ortoflavivirus. Se generaron cepas infecciosas de ZIKV, YFV y el virus Langat (LGTV; un ortoflavivirus transmitido por garrapatas) [ 63 ], y se infectaron células humanas con una MOI de 0,1 en presencia de concentraciones crecientes de DMA-177 o DMA-176 ( Fig. 6D y 6E ). El DMA-176 inhibió la replicación del ZIKV, pero no mostró una inhibición significativa del YFV ni del LGTV. Por el contrario, el DMA-177 inhibió la replicación del ZIKV, el YFV y el LGTV, lo que indica una actividad antiviral más amplia frente a los ortoflavivirus. El diferente efecto observado con el DMA-176 y el DMA-177 sugiere que sutiles diferencias estructurales entre los SLA influyen en la sensibilidad del compuesto. En conjunto, los resultados muestran que las moléculas pequeñas dirigidas al SLA pueden inhibir la replicación de los ortoflavivirus, lo que representa una prueba de concepto para una amplia actividad antiviral.
Discusión
Aquí presentamos evidencia que respalda la conservación funcional del promotor de ARN SLA entre genomas de ortoflavivirus patógenos. La capacidad del SLA de diferentes ortoflavivirus transmitidos por mosquitos y garrapatas para sustentar la replicación del ARN del DENV y el ZIKV, junto con características estructurales y de secuencia conservadas, establece un modo funcional común de reconocimiento del promotor NS5 por todos estos patógenos virales humanos. Utilizando virus recombinantes, junto con modelado estructural y simulaciones de dinámica molecular, definimos relaciones estructura-función compartidas y específicas de virus de los subelementos SLA en ortoflavivirus. Nuestros datos respaldan que la divergencia de los elementos estructurales SLA en virus de diferentes grupos ecológicos coevolucionó para sustentar un mecanismo común para la replicación viral. Con base en esta universalidad de los SLA, examinamos una biblioteca de moléculas pequeñas centrada en el ARN en busca de ligandos del SLA del DENV y encontramos moléculas que muestran inhibición no solo del DENV sino también de otros ortoflavivirus patógenos.
Dirigirse a las estructuras de ARN virales representa una estrategia antiviral emergente y prometedora, pero su éxito depende de una comprensión detallada de la función y estructura del ARN [ 64 ]. Aquí encontramos dos moléculas de DMA como ligandos de SLA que inhiben la replicación de diferentes ortoflavivirus. Se ha informado previamente que los DMA se unen a estructuras de ARN específicas de diferentes virus, incluidos el VIH, el enterovirus A71 y el SARS CoV2 [ 65 – 68 ] mostrando potencial como fármacos antivirales. Se ha demostrado que DMA-135 se une al SLII del IRES del enterovirus-A71 [ 67 ]. Estudios estructurales y biofísicos respaldaron un mecanismo alostérico en el que DMA-135 induce un cambio conformacional en el ARN SLII que estabiliza un complejo ternario con la proteína AUF1 [ 68 ] . Encontramos DMA-176 y DMA-177 como ligandos de DENV SLA. DMA-177 mostró inhibición de miembros de MBFV y TBFV, lo que proporciona una prueba de concepto para un efecto de amplio espectro sobre la infección por ortoflavivirus. En contraste, DMA-176 inhibió DENV y ZIKV, sin un efecto significativo sobre la replicación de YFV o LGTV. El impacto diferencial de DMA-176 y DMA-177 podría explicarse por las distintas propiedades estructurales de las SLA de YFV y TBFV en comparación con las de otros MBFV ( Figs. 3 y 4 ). No obstante, serán necesarios más estudios bioquímicos y estructurales para definir el mecanismo de acción de estos DMA sobre la replicación de ortoflavivirus.
En consonancia con nuestros hallazgos, estudios previos han informado que los SLA de ciertos ortoflavivirus pueden ser intercambiables [ 40 , 46 , 54 , 69 ]. Sin embargo, se ha descrito una falta de compatibilidad de SLA entre JEV y WNV, dos miembros del grupo MBFV [ 54 ]. Aunque inicialmente desconcertante, esta observación puede explicarse por el estricto requisito del dominio de metiltransferasa (MTasa) NS5 de WNV para nucleótidos específicos en las posiciones dos y tres del genoma viral (5′-AGU), mientras que JEV contiene un motivo 5′-AGA en estas posiciones [ 53 ]. De acuerdo con esta interpretación, aquí encontramos que los SLA de MBFV cuyos genomas portan naturalmente un motivo 5′-AGA, incluyendo SLEV, JEV, ILHV, USUV y ROCV, no son funcionales en el contexto de DENV, que, al igual que WNV, contiene una secuencia 5′-AGU ( Fig. S1 ). Es importante destacar que la sustitución de la tercera posición de adenina por uridina en estos SLA fue suficiente para restaurar la replicación viral en todos los virus quiméricos MBFV ( Fig. S1 ). Curiosamente, los SLA de TBFV, que todos se inician con el motivo 5’AGA, son funcionales en el contexto de quimeras de DENV y ZIKV. En línea con esta observación, se ha informado previamente que WNV NS5 es competente para la metilación N-7 in vitro del ARN de POWV (un TBFV) [ 53 ]. Esto indica que la secuencia 5′-AGA puede integrarse cuando se presenta dentro del contexto estructural de TBFV SLA. En conjunto, las observaciones previas y la aquí presentada respaldan la compatibilidad funcional de la secuencia completa de TBFV SLA en el contexto de MBFV.
A diferencia de la compatibilidad funcional observada entre MBFV y TBFV, el SLA del c-ISFV AeFV no logró sustentar la replicación de DENV o ZIKV en células de vertebrados o insectos y mostró una unión reducida a DENV NS5, lo que indica una divergencia de la interfaz NS5-SLA más allá de la compatibilidad funcional. El SLA del dhl-ISFV NHUV, que es filogenéticamente más cercano a los MBFV, conservó la capacidad de unirse a DENV NS5 con una afinidad aparente comparable a la del SLA homólogo de DENV ( Fig. 5A ). La proximidad filogenética, reflejada en características estructurales similares, probablemente explica por qué se pudo lograr la compatibilidad funcional después de una adaptación limitada en el cultivo celular. En este sentido, la replicación del virus quimérico NHUV SLA se asoció con la selección de una sola mutación en el TS. Aunque el mecanismo molecular por el cual esta mutación permite la replicación productiva aún no está claro, la tolerancia a las protuberancias y los desajustes en el TS de los ortoflavivirus sugiere que la estabilidad y la geometría local del tallo, en lugar de la secuencia primaria, pueden ser relevantes para la competencia del promotor.
Los análisis funcionales, estructurales y computacionales revelan la divergencia de los subelementos SLA de los SLA de MB y TBFV. Las diferencias en la secuencia 3WJ y las longitudes de las uniones, así como en la longitud y composición del SSL, distinguen a los SLA de estos dos grupos de virus. Dentro del mismo grupo viral, los 3WJ están conservados ( Fig. 4A ), lo que sugiere posibles interacciones con factores específicos del huésped. Estudios previos han propuesto que el 3WJ desempeña un papel clave en la configuración de la conformación 3D del SLA [ 43 , 44 ]. Los análisis de RMN indican que el 3WJ es parcialmente flexible, y estudios estructurales adicionales han propuesto que esta flexibilidad permite al SLA adoptar distintas arquitecturas globales. Por lo tanto, el 3WJ puede proporcionar plasticidad de movimiento al SLA para la transición entre diferentes etapas en el proceso de replicación del ARN viral.
A pesar de la divergencia entre los subelementos SLA, varias características clave permanecen conservadas en los MBFV y TBFV, incluyendo el apilamiento coaxial conservado del SS con el TS y la variación de secuencia restringida dentro del TL ( Fig. 3 ). Aunque el TL de cuatro nucleótidos podría generar teóricamente 256 combinaciones de secuencia, solo se observan ocho en la naturaleza, lo que refleja fuertes restricciones funcionales. Los contactos específicos entre los nucleótidos del TL y los residuos conservados de RdRp, junto con una distancia espacial conservada entre el TL y el extremo 5′, respaldan aún más el modo de unión NS5 compartido en estos virus. El SLA del YFV muestra un 3WJ único y la distancia medida entre el TL y el extremo 5′ es mayor que la observada para otros MBFV y TBFV ( Fig. 4H ). La posición divergente del YFV dentro del grupo MBFV es consistente con las propiedades estructurales únicas del SLA de este virus. Sin embargo, se encontró que el SLA del YFV es funcional en el contexto del DENV y el ZIKV.
En conjunto, nuestros datos indican que la replicación viral productiva requiere no solo estructuras conservadas y secuencias específicas dentro de los subelementos SLA individuales, sino también su organización 3D coordinada. Aunque ciertas características de SLA han divergido de manera específica de grupo, la combinación de estos elementos mantiene una arquitectura global conservada en los SLA de ortovirus patógenos que permite el reconocimiento heterólogo de NS5 y la replicación del ARN viral. Estudios previos han demostrado que la unión de NS5 a SLA induce alteraciones en el patrón de huella de ARN en el 3′ del BS, lo que respalda cambios conformacionales en el ARN lejos de los sitios de unión de NS5 propuestos [ 46 ]. Estudios más recientes también sugirieron que, tras la formación del complejo RNP, tanto los dominios NS5 como el SSL del SLA experimentan un reordenamiento conformacional [ 49 , 70 ], lo que resalta la complejidad de la interacción SLA-NS5. La falta de información estructural que permita visualizar el complejo SLA-NS5 con el extremo 3′ del ARN acoplado al canal de la plantilla de la polimerasa, lo que podría requerir reordenamientos estructurales de NS5, representa un desafío clave para comprender mejor el mecanismo de iniciación de la síntesis de ARN viral.
Se ha propuesto que la unión de los dominios RdRp y MTasa de NS5 al extremo TL y 5′ del SLA tiene relevancia funcional [ 43 , 45 ] y se asignó un papel crítico a la región de enlace entre estos dos dominios [ 71 ]. También se ha informado de la unión cooperativa de RdRp y MTasa al SLA [ 41 , 43 , 49 , 72 ]. Sin embargo, durante la infección, las dos actividades enzimáticas actúan sobre diferentes moléculas de ARN viral, por lo que aún no está claro el vínculo entre estas dos actividades. En este sentido, la síntesis de ARN viral de cadena negativa y la metilación del ARN viral son dos eventos que tienen lugar en diferentes etapas del proceso de replicación del ARN viral. Mientras que la síntesis de la cadena negativa se inicia en el extremo 3′ del ARN genómico de la cadena positiva, utilizando un promotor SLA que ya está capuchado y metilado, el proceso de metilación del capuchón tiene lugar en el extremo 5′ de una molécula de ARN de cadena positiva recién sintetizada. En este sentido, el mecanismo de iniciación de la síntesis de ARN de cadena positiva aún no está claro. Recientemente se ha informado que el extremo 3′ de la cadena negativa forma una estructura similar a SLA (SLA’) que se une a NS5 pero es inactiva para la iniciación de la síntesis de ARN de cadena positiva [ 59 ]. Es posible que el SLA de la cadena positiva también sirva como promotor para la síntesis de nueva cadena positiva mediante un proceso de iniciación trans. La iniciación trans in vitro por DENV2 NS5 se ha observado previamente, utilizando como promotor un SLA de una molécula para iniciar en el extremo 3′ de una molécula diferente [ 31 , 33 , 36 ]. En este caso, la unión de NS5 a una SLA recién sintetizada facilitaría la metilación de la caperuza. Concluimos que la replicación del ARN viral en células infectadas requiere múltiples interacciones coordinadas entre SLA y NS5, y que varios aspectos mecanísticos clave de este proceso aún no se han resuelto.
Más allá de las actividades de RdRp y MTasa, la NS5 de los ortoflavivirus participa en la evasión inmunitaria del huésped [ 73 ]. Por ejemplo, en el caso del DENV, la NS5 media la degradación de STAT2 y ERC1 [ 74 , 75 ]. Se espera que los diferentes entornos inmunitarios innatos en distintos huéspedes impongan distintas presiones selectivas, lo que conduce a la divergencia evolutiva de la NS5 entre los grupos ecológicos. Si bien los sitios de unión de la NS5 a SLA son esenciales para la replicación viral de todos los miembros del género, las regiones de la NS5 que median la evasión inmunitaria pueden experimentar una divergencia evolutiva impulsada por el huésped. Esta divergencia en la secuencia y estructura de la NS5 podría modular las conformaciones de la proteína, lo que, a su vez, podría afectar el reconocimiento de SLA en distintos grupos ecológicos.
Dado que SLA se encuentra dentro de la región 5’UTR, no está limitado por los requisitos de codificación de proteínas y, por lo tanto, puede explorar un panorama estructural y evolutivo más amplio. Proponemos que la mayor parte de la divergencia ecológica de secuencia y estructura de SLA observada surge de la coevolución con NS5 bajo presiones selectivas específicas del huésped. Sin embargo, no excluimos la posibilidad de que ciertas características de SLA, como la divergencia de 3WJ, también reflejen adaptaciones evolutivas impulsadas por restricciones ecológicas o funcionales, contribuyendo a la diversificación de SLA en los grupos de ortoflavivirus. Los subelementos conservados ecológicamente dentro de SLA pueden actuar como firmas moleculares para ortoflavivirus patógenos humanos, proporcionando una herramienta para la detección temprana de virus patógenos emergentes en reservorios naturales.
En resumen, nuestro trabajo define propiedades estructurales y funcionales clave de un elemento esencial del ARN viral conservado entre los ortoflavivirus patógenos. La identificación de pequeñas moléculas que se unen a SLA e inhiben la infección de múltiples virus ofrece una oportunidad para desarrollar estrategias antivirales contra ortoflavivirus conocidos y emergentes.
Materiales y métodos
Bases de datos
A partir de la página web del ICTV sobre el género Orthoflavivirus , creamos una base de datos que contiene 92 virus ( Tabla S1 ). Dentro de estos virus se incluyen las especies de orthoflavivirus y las especies relacionadas no clasificadas, con excepción de los virus flavi-like segmentados. Además, creamos una base de datos SLA seleccionando aquellos virus cuyas secuencias 5′ comienzan en AG, que consiste en SLA de 38 virus ( Tabla S2 ).
Alineamientos de secuencias
Para el alineamiento de secuencias de ARN se utilizó Clustal Omega [ 76 ], mientras que el alineamiento de secuencias de proteínas NS5 se obtuvo utilizando MUSCLE [ 77 ]. Los logotipos de secuencias se obtuvieron utilizando el servidor web WebLogo3 [ 78 ]. A partir del alineamiento de secuencias de proteínas NS5 construimos árboles filogenéticos utilizando los servidores web IQ-TREE [ 79 ] e iTOL [ 80 ].
Predicción y análisis de la estructura 2D mediante SLA
La estructura secundaria SLA está disponible a partir de datos experimentales solo para DENV1, DENV2, DENV3, DENV4, ZIKV, WNV y YFV [ 47 , 48 , 81 – 83 ]. Para los demás virus, la estructura secundaria SLA se predijo a través del servidor web RNAfold de la suite web Vienna RNA [ 84 ] utilizando el modelo de Turner 1999 [ 85 ]. El servidor web y el modelo se eligieron debido a la similitud entre las estructuras secundarias predichas y experimentales de DENV1 a 4, ZIKV y YFV. Forna se utilizó como herramienta para la visualización de ARN 2D [ 86 ].
Alineaciones considerando estructuras 2D
Para alinear múltiples secuencias de ARN y predecir una estructura secundaria de consenso, utilizamos LocRNA 2.0 [ 87 ]. Como entrada, empleamos la secuencia y las estructuras 2D predichas mediante el servidor web RNAfold con el modelo de Turner de 1999. Se calculó la conservación de la estructura secundaria para cada posición en la alineación. Evaluamos si el nucleótido estaba ausente, si se encontraba en cadena simple o en cadena doble, y calculamos un índice de conservación, definido como la frecuencia con la que el estado de esa posición coincidía con la estructura de consenso.
Análisis de la estructura 2D mediante SLA
A partir de las estructuras 2D y la alineación, se determinaron los siguientes parámetros para cada SLA:
La longitud del tallo se consideró a partir del número de nucleótidos de cadena simple o doble en el extremo 5′ del tallo.
La secuencia TL de 4 nucleótidos se determinó a partir de una alineación de secuencias TL. Se utilizaron las secuencias TL de DENV2 y ZIKV como referencia para definir secuencias TL de 4 nucleótidos de longitud para cada virus.
Los nucleótidos en el 3WJ se identificaron en función de su alineación con las regiones de bucle del 3WJ en la estructura de consenso obtenida a partir de alineaciones de secuencia-estructura específicas del grupo.
Dinámica molecular
El modelo atómico para el complejo RNP SLA-NS5 se derivó de la estructura Cryo-EM con PDB ID 8GZP. Las regiones internas que no se resolvieron debido a su alta flexibilidad se modelaron por homología utilizando el programa SWISS-MODEL [ 88 ] basado en la secuencia NS5-DENV3 reportada en la estructura. De manera similar, para el SLA (nucleótidos 1–69), dos nucleótidos del bucle lateral del tallo no se resolvieron y se modelaron utilizando ModeRNA [ 89 ], un programa de modelado por homología específico para ARN. Aunque la estructura se resolvió en presencia de la caperuza 5′, la guanosina difosfato y un catión Mg²⁺ en el sitio catalítico de la MTasa, ambos se eliminaron debido a la evidencia experimental que muestra que su presencia no afecta la interacción SLA-NS5 [ 90 ]. Los dos cationes Zn²⁺ resueltos se incluyeron en la estructura.
La proteína y el ARN se modelaron utilizando el campo de fuerza Amber ff14SB [ 91 ] con la corrección ILDN para residuos de proteína y parámetros diedros actualizados para el ARN [ 92 ]. Los parámetros para los cuatro aminoácidos (C778, C847, H712 y H714) que forman un entorno tetraédrico de Zn2+ se obtuvieron del campo de fuerza Zinc AMBER (ZAFF) [ 93 ]. Los parámetros para los otros cuatro aminoácidos (C446, C449, H441 y E437) que coordinan el segundo Zn2+ se derivaron utilizando el Metal Center Parameter Builder en Python: MCPB.py [ 94 ]. Las interacciones modeladas por estos parámetros actúan como enlaces de tipo covalente entre el metal y los cuatro aminoácidos coordinantes. Para obtener los últimos parámetros de Zn2+, primero se calcularon las distancias y ángulos de equilibrio entre átomos para el sistema en fase gaseosa utilizando métodos ab initio. A continuación, se obtuvieron las constantes de fuerza mediante la minimización de un modelo aproximado del sistema sin el metal (por ejemplo, convirtiendo el grupo tiol (SH) en tiolato (SO⁻) en los residuos de cisteína que coordinan el metal), utilizando la teoría del funcional de la densidad (DFT) al nivel B3LYP/6-31G*. Finalmente, se asignaron cargas parciales mediante el método RESP, implementado en Gaussian 16.
Los estados de protonación de los residuos ionizables se determinaron en función de los valores de pKa estimados utilizando PropKa 3.0 [ 95 ], un predictor empírico de pKa para residuos de superficie y enterrados. Después de inspeccionar el entorno de cada histidina, se asignaron como HID: H215, H263, H441, H512, H712, H714 y H768; el resto se asignaron como HIE. Utilizando el módulo tLeap [ 96 ] el sistema se solvató utilizando aproximadamente 45 000 moléculas de agua TIP3P [ 97 ] colocadas en una caja octaédrica de 15 Å de la superficie de la proteína. Se obtuvo una concentración de sal fisiológica de 0,15 M añadiendo 123 iones Na+ y Cl-. Se incluyeron 53 Na+ adicionales para neutralizar la carga del sistema (-53 e).
Se realizaron simulaciones por triplicado utilizando el módulo PMEMD Cuda del paquete Amber 22 [ 96 ]. Cada réplica incluyó las tres etapas de preproducción (minimización, calentamiento y equilibración). Al comienzo de la preproducción, las moléculas de agua y los iones se minimizaron energéticamente con restricciones posicionales (300 kcal/mol/Ų) en los átomos de biomoléculas (proteínas y ARN). Esto implicó 500 pasos de descenso más pronunciado seguidos de 500 pasos de gradiente conjugado. A continuación, se eliminaron las restricciones y se realizaron 500 pasos más de descenso más pronunciado y 2500 pasos de gradiente conjugado. Tras la minimización de energía, el sistema se calentó a 300 K y 1 bar en tres etapas, todas a volumen constante (conjunto NVT): primero, la temperatura se incrementó de 0 a 100 K durante 5 ps con fuertes restricciones (100 kcal/mol/Ų) en los átomos de la biomolécula, relajando solo el disolvente y los iones; segundo, las restricciones se redujeron a 25 kcal/mol/Ų y la temperatura se elevó a 200 K durante 10 ps; y tercero, la temperatura se incrementó de 200 K a 300 K con restricciones de 10 kcal/mol/Ų durante 100 ps. Finalmente, el equilibrio a presión constante se realizó en dos pasos (conjunto NPT): primero, 100 ps a 300 K y 1 bar con restricciones de 10 kcal/mol/Ų; y segundo, 10 000 ps a 300 K y 1 bar sin restricciones. Se utilizó un paso de tiempo de 1 fs durante el calentamiento; 2 fs durante el equilibrio. La temperatura se controló con el termostato de Langevin (frecuencia de colisión: 1 ps ⁻ ¹) [ 98 ], y la presión, para el equilibrio NPT, se mantuvo con el barostato de Berendsen (tiempo de relajación: 2 ps) [ 99 ]. El algoritmo SHAKE [ 100 ] restringió los enlaces que involucran hidrógenos no polares y se utilizó un corte de 10 Å para las interacciones no enlazadas. Se realizaron tres ejecuciones de producción independientes de 2 μs para cada sistema utilizando el conjunto NVT y un paso de tiempo de 2 fs. Los primeros 0,5 μs de cada trayectoria se descartaron como equilibrio adicional.
Predicción de estructura 3D SLA
La predicción 3D se realizó utilizando RNAComposer [ 101 ]. Se utilizó la predicción secundaria y se obtuvieron 5 modelos diferentes para cada SLA. Para visualizar las estructuras 3D, se utilizaron VMD y ChimeraX [ 102 , 103 ].
Análisis de la estructura 3D mediante SLA
Utilizamos DSSR [ 104 ] para analizar las estructuras 3D. Las distancias de los extremos TL-5′ se calcularon utilizando el origen del marco de referencia para las dos bases, tal como se define en el software DSSR. La segunda posición de la TL se eligió como referencia de la TL, identificada mediante la división en pisos de la longitud de la horquilla TS y la resta de 1 para cada SLA.
El valor RMSD de TL se calculó para cada modelo 3D con respecto a la estructura DV4 unida a NS5 (pdb 8gzp). Todos los valores RMSD se calcularon después de alinear TL con la misma estructura de referencia y solo se incluyeron los átomos del esqueleto del ácido nucleico (OP2, OP1, P, O5′, C5′, C4′, C3′, O3′).
Se identificó el apilamiento coaxial con DSSR como una hélice de al menos 10 pares de bases, con el primer y último par separados por no más de 8 nucleótidos (criterios generosos para TL o SSL). El apilamiento coaxial del tallo lateral (SS) con el tallo superior (TS) observado en las estructuras del complejo SLA-NS5 es estable a lo largo de las tres simulaciones de dinámica molecular de microsegundos independientes (97 % de las conformaciones).
Construcción de virus quiméricos y subquiméricos
Para construir los virus quiméricos que contienen mutaciones en el SLA, modificamos clones infecciosos de DENV2 [ 51 , 105 ] y ZIKV [ 50 ]. Se introdujeron mutaciones en el ICRep DENV2 reemplazando el fragmento MluI-NotI con los fragmentos respectivos derivados de PCR ( Tabla S3 ) o PCR superpuestas ( Tabla S4 ) que contenían la secuencia deseada. En el caso del ZIKV, se introdujeron mutaciones en el IC ZIKV reemplazando el fragmento MluI-XhoI y en el ICRep ZIKV reemplazando el fragmento MluI-AvrII con fragmentos derivados de PCR ( Tabla S3 ) o PCR superpuestas ( Tabla S4 ) que contenían la secuencia deseada. La Tabla S5 contiene la secuencia de los cebadores utilizados.
Los productos de ligación se transformaron en bacterias XL1-Blue y se obtuvieron varios clones para cada mutante. Los plásmidos resultantes se secuenciaron y los clones positivos se utilizaron para la transcripción de ARN in vitro.
Transcripción y transfección in vitro
Los plásmidos que contenían los virus de tipo salvaje y quiméricos se linealizaron con XbaI para DENV2 y KpnI para ZIKV; y se utilizaron para la transcripción in vitro con la ARN polimerasa T7 (Ambion) en presencia del análogo de cap m7GpppA (NEB). Modificamos los dos últimos nucleótidos del promotor T7 (GG por GA) para que los transcritos se iniciaran con el auténtico m7GpppAGU. La cuantificación del ARN se evaluó con un Qubit4 (Invitrogen) y la integridad del ARN se confirmó en geles de agarosa al 1%. Los transcritos de ARN se transfectaron en células BHK y C6/36HT utilizando Lipofectamine 2000 y medio Opti-MEM (Invitrogen). Para placas de cultivo celular de 24 pocillos, se utilizaron 50 ng de ARN viral por pocillo.
cultivo celular
Las transfecciones e infecciones se realizaron utilizando las siguientes líneas celulares. Las células C6/36HT (ATCC CRL-1660), una línea celular de Aedes albopictus adaptada para crecer a 33 °C, se cultivaron en medio Leibovitz L-15 suplementado con 10 % de suero fetal bovino (FBS), 100 U/ml de penicilina, 100 μg/ml de estreptomicina, 0,3 % de caldo de triptosa fosfato, 0,02 % de glutamina, 1 % de solución de aminoácidos no esenciales del medio mínimo esencial (MEM) y 0,25 μg/ml de anfotericina B (Fungizone). Las células de riñón de hámster bebé (BHK, ATCC, CCL-10) se cultivaron en medio mínimo esencial α (MEMα) (Gibco, Thermo Fisher Scientific) suplementado con 10 % de suero fetal bovino (FBS) (Gibco, Thermo Fisher Scientific) y 100 U/ml de penicilina-estreptomicina en placas de cultivo celular de 100 mm y se incubaron a 37 °C con 5 % de CO2. Las células A549 (línea celular epitelial de adenocarcinoma de pulmón humano, ATCC, CCL-185) se cultivaron en medio Eagle modificado de Dulbecco con mezcla de nutrientes F-12 (DMEM-F-12) suplementado con 10 % de suero fetal bovino, 100 U/ml de penicilina y 100 μg/ml de estreptomicina.
Actividad de la luciferasa y análisis estadístico de la replicación de DENVRep.
Los ensayos de luciferasa de Renilla se realizaron utilizando el kit del sistema de ensayo de luciferasa de Renilla (Promega). La replicación de los virus reporteros se clasificó integrando el comportamiento temporal de la señal de luciferasa y comparándola con el control no replicativo (NS5 mut).
Los virus quiméricos se clasificaron como replicativos, subóptimos y no replicativos. Se definió una quimera como replicativa cuando la señal de luminiscencia no difería significativamente del tipo silvestre en cada punto temporal, como subóptima cuando se detectó un aumento estadísticamente significativo de la señal en relación con el control no replicativo a las 72 horas posteriores a la transfección, y como no replicativa cuando la señal de luciferasa no difería significativamente del control no replicativo. Las comparaciones estadísticas se realizaron mediante pruebas t de Welch de dos colas, teniendo en cuenta tres réplicas biológicas.
Ensayo de inmunofluorescencia
Las células BHK-21 se cultivaron en placas de cultivo celular de 24 pocillos que contenían cubreobjetos de vidrio de 12 mm. En los tiempos correspondientes después de la transfección o infección, se recogieron los cubreobjetos y las células se fijaron con metanol durante 15 min a –20 °C. Los cubreobjetos se bloquearon con gelatina al 0,2 % (Sigma) en PBS y se incubaron con anticuerpo monoclonal de ratón anti-E (E18) diluido 1/500 en solución de bloqueo para DENV2 y con anticuerpo policlonal de conejo anti-NS3 diluido 1/500 en solución de bloqueo para ZIKV. Se empleó anticuerpo de cabra anti-conejo conjugado con Alexa Fluor 488 (Thermo Fisher Scientific) para detectar el anticuerpo primario y se añadió DAPI (Thermo Fisher Scientific) para visualizar los núcleos. Las imágenes se obtuvieron utilizando un microscopio de fluorescencia invertido Axio Observer 3 (Zeiss), con objetivo de 10x.
Retrotranscripción
Se extrajo el ARN viral de los sobrenadantes de las células transfectadas utilizando TRIzol (Invitrogen). El ARN purificado se utilizó para realizar reacciones de transcripción inversa-PCR (RT-PCR) por duplicado con cebadores diseñados específicamente para amplificar los primeros 500 nucleótidos de los extremos del genoma de ZIKV y DENV2, utilizando la transcriptasa inversa SuperScript III (Invitrogen) y la ADN polimerasa Platinum Pfx (Invitrogen).
RT-qPCR
Para la qRT-PCR del ARN viral, se utilizaron muestras de ARN del sobrenadante para la transcripción inversa, como se describió previamente. Las reacciones se realizaron por duplicado en placas de 96 pocillos utilizando 2 μl de la mezcla de reacción de RT como plantilla, 5 μl de FastStart SYBR green Master 2 × mix (Roche), una concentración de 300 nM de cada cebador y agua libre de RNasa hasta un volumen final de 10 μl. Para el DENV2, se utilizaron los cebadores AVG1117 (5′ACAAGTCGAACAACCTGGTCCAT3′) y AVG1118 (5′GCCGCACCATTGGTCTTCTC3′) para amplificar los nucleótidos 9937 a 10113 dentro de la secuencia codificante de NS5. Para el ZIKV, se utilizaron los cebadores AVG2098 (5′GCCGCCACCAAGATGAACTGATTG3′) y AVG2099 (5′GCAGTCTCCCGGATGCTCCATC3′) para amplificar los nucleótidos 9854 a 9927 dentro de la secuencia codificante NS5.
Ensayos de desplazamiento de movilidad electroforética
Para obtener ARN, primero se preparó una plantilla de ADN mediante amplificación por PCR. Las secuencias de nucleótidos de los cebadores utilizados y las plantillas para la PCR de cada ARN sintético se detallan en la Tabla S6 . A continuación, se realizó la transcripción in vitro de las plantillas utilizando la ARN polimerasa T7 (180 min, 37 °C) y se trataron con DNasa I libre de RNasa para eliminar las plantillas. Los productos de ARN se purificaron mediante electroforesis en gel de poliacrilamida nativa, seguida de la escisión de las bandas correspondientes y la elución. La cuantificación del producto final se evaluó utilizando un fluorómetro Qubit4 (Invitrogen), y la integridad se confirmó en geles de agarosa al 2 %. Las secuencias de ARN sintético se especifican en la Tabla S6 .
Para la expresión de proteínas, se utilizó un plásmido pQE-30 que contenía la secuencia de la proteína NS5 completa, como se usó anteriormente [ 90 ]. La NS5 se expresó en Escherichia coli Rosetta (pLacI) durante la noche a 18 °C después de la inducción con 100 μM de isopropil-β-d-tiogalactopiranósido (IPTG). La lisis de las células se llevó a cabo mediante sonicación en tampón de unión (50 mM de fosfato de sodio, pH 7,5, 600 mM de NaCl, 5 mM de MgCl2 y 10 % de glicerol) en presencia de 0,2 % v/v de β-mercaptoetanol, 1 % de Triton X-100, 12 µg/ml de DNasa I, 1 mM de PMSF y 1 % v/v de cóctel de inhibidores de proteasa (Sigma). Después de la centrifugación, el sobrenadante se cargó en una columna de afinidad His-Trap nickel Sepharose (GE Healthcare) y se lavó dos veces con tampón de unión más 80 mM de imidazol, y las proteínas se eluyeron con tampón de unión que contenía 300 mM de imidazol. El tampón de muestra se cambió a tampón de HPLC (tampón Tris-HCl 10 mM, pH 7,5, 300 mM NaCl, 5 mM MgCl 2 , 10% glicerol y 1 mM ditiotreitol) usando filtros centrífugos Amicon Ultra, 10 kDa MWCO (Merck Millipore). La proteína se purificó aún más por cromatografía de exclusión por tamaño usando una columna Superdex 200 (Cytiva). La proteína se concentró usando un filtro centrífugo Amicon Ultra, 50 kDa MWCO (Merck Millipore) y se almacenó a −20 °C en tampón de HPLC que contenía 40% de glicerol.
Para EMSA, la mezcla de reacción de unión contenía 5 mM de HEPES (pH 7,9), 20 mM de NaCl, 2 mM de MgCl₂ , 3,8 % de glicerol, 10 nM de sonda de ARN y concentraciones crecientes de la proteína. Los complejos ARN-NS5 se analizaron mediante electroforesis en geles de poliacrilamida al 5 % nativos suplementados con 5 % de glicerol. Los geles se precorrieron durante 10 min a 80 V, luego se cargaron 20 μl de muestra y se dejó que la electroforesis procediera durante 1 h a 4 °C a un voltaje constante de 60 V. Los geles se tiñeron con SYBR Gold (Thermo Fisher Scientific) y se visualizaron con un transiluminador.
Detección de ligandos de ARN
Determinación de la puntuación Z. El ARN se hibridó a 95 °C durante 5 minutos y se enfrió rápidamente en hielo durante 30 minutos. Después de la hibridación del ARN, se preparó una solución de DENV SLA 0,085 μM y TO-PRO-1 0,5 μM en tampón (5 mM HEPES (pH 7,9), 25 mM KCl, 2 mM MgCl2) y se incubó en un agitador durante 30 minutos. Se añadieron 5 μL de neomicina 10 o 25 μM o tampón, seguidos de 5 μL de la solución de ARN-colorante, a una placa negra de 384 pocillos. La placa se centrifugó durante 1 minuto y se dejó incubar durante 30 minutos. Después de la incubación, la placa se leyó en un lector de placas CLARIOstar (BMG Labtech) (excitación = 492 nm; emisión = 575). La puntuación Z’ se calculó según los estándares de la literatura [ 106 ].
Cribado de moléculas pequeñas. El ARN se hibridó a 95 °C durante 5 minutos y se enfrió rápidamente en hielo durante 30 minutos. Se sembraron 10 nL de moléculas pequeñas en placas negras de 384 pocillos utilizando un manipulador de líquidos acústico Echo 550 (Labcyte). Se prepararon soluciones de DENV SLA de 0,085 μM y TO-PRO-1 de 0,5 μM utilizando un tampón (5 mM HEPES (pH 7,9), 25 mM KCl, 2 mM MgCl2) y luego las soluciones se agitaron y se incubaron durante 30 minutos. Se transfirieron 10 μL de la solución de ARN-colorante a la placa utilizando un dispensador de líquidos ThermoFisher y luego las placas se agitaron, se centrifugaron y se dejaron incubar durante 30 minutos. Tras la incubación, la placa se leyó en un lector de placas CLARIOstar (BMG Labtech) (excitación = 492 nm; emisión = 575 nm). El porcentaje de desplazamiento se calculó mediante la siguiente ecuación, donde F es la fluorescencia del complejo ARN-colorante con moléculas pequeñas y F0 es la fluorescencia del complejo ARN-colorante sin moléculas pequeñas. El cribado se realizó por triplicado. Se consideraron positivos los compuestos que mostraron una FID mayor o igual al 20 % en al menos 2/3 de las réplicas y que presentaron una FID promedio mayor o igual al 20 % en las tres réplicas.
acciones virales
Para obtener cepas virales de DENV2 y ZIKV, se recolectaron los sobrenadantes de células C6/36 HT transfectadas a los 7 días posteriores a la transfección (dpt), se centrifugaron a 1000 rpm durante 5 minutos y posteriormente se titularon. Para YFV, se transfectó ARN correspondiente al virus 17D [ 107 ] en células BHK y se recolectó el sobrenadante a los 3 dpt, se centrifugó a 1000 rpm durante 5 minutos y posteriormente se tituló. Finalmente, para LGTV, se transfectó ARN correspondiente al virus TP21 [ 63 ] en células BHK y se recolectó el sobrenadante a los 3 dpt, se centrifugó a 1000 rpm durante 5 minutos y posteriormente se tituló.
Ensayos de inhibición de la infección
Las células A549 se infectaron con cepas virales diluidas en medio F-12. Las infecciones por DENV2 y LGTV se realizaron con una multiplicidad de infección (MOI) de 0,1, mientras que las infecciones por YFV y ZIKV se llevaron a cabo con una MOI de 0,01. Una hora después de la infección (hpi), se retiró el inóculo y se reemplazó con medio F-12 que contenía la concentración indicada de la molécula pequeña, suplementado con 1 % de DMSO y 2 % de suero fetal bovino. Los cultivos infectados se recolectaron a las 24 hpi.
Ensayo de viabilidad celular
La viabilidad de las células A549 tras el tratamiento con inhibidores se determinó midiendo los niveles de ATP celular, un indicador de la actividad metabólica. Para ello, se utilizó el kit CellTiter-Glo Luminescent Cell Viability Assay (Promega), siguiendo las instrucciones del fabricante.
ensayos de placa
Las células BHK-21 se sembraron en placas de cultivo celular de 24 pocillos y se cultivaron durante la noche. Los sobrenadantes de las células BHK-21 transfectadas o infectadas se diluyeron en serie y se utilizaron 200 µl del inóculo para infectar monocapas de BHK-21 durante 1 hora a 37 °C. Posteriormente, se añadió 1 ml de medio de recubrimiento (MEMα, 0,8 % de metilcelulosa, suplementado con 5 % de FBS) a cada pocillo. Las células se fijaron 7 días después de la infección (dpi) con formaldehído al 10 % y se tiñeron con cristal violeta.
Información complementaria
Figura S1. Impacto de la identidad del tercer nucleótido del genoma viral en la replicación del ARN de quimeras DENVRep que portan SLA de diferentes MBFV.
Se muestra la replicación de quimeras DENVRep que portan SLA de SLEV, JEV, ILHV, USUV y ROCV. En cada caso, se indican los virus quiméricos que contienen la A nativa en la tercera posición (5’AGA) o U (5’AGU). Actividad de luciferasa normalizada en función del tiempo en células BHK transfectadas con ARN correspondientes a DENVRep WT, quimeras y NS5 Mut, como se indica en la parte superior. La comparación entre cada quimera que porta 5’AGA o 5’AGU a las 72 h se realizó mediante pruebas t de Welch de dos colas. * indica un valor p ajustado <0,05.
https://doi.org/10.1371/journal.ppat.1014233.s001
(EPS)
Figura S2. Interacciones entre G32 del bucle superior de SLA y R770 (azul) o K841 (naranja) de la región del pulgar de NS5 RdRp.
Distribuciones de densidad de probabilidad de las distancias calculadas a partir de trayectorias de dinámica molecular agregadas (3 μs en total de tres réplicas). Se muestran estimaciones de densidad de kernel para cada interacción (ancho de banda determinado mediante la regla de Scott). Las líneas verticales discontinuas indican las distancias correspondientes en la estructura inicial (PDB 8gzp), resaltando las desviaciones de la geometría de contacto inicial durante las simulaciones. Las distancias se calcularon utilizando el oxígeno carbonílico (O6) de la base de guanina (G32), el átomo de carbono central (CZ) del grupo guanidinio R770 y el átomo de nitrógeno (NZ) del grupo ε-amonio de K841. Los datos se visualizaron utilizando Seaborn (1).
https://doi.org/10.1371/journal.ppat.1014233.s002
(EPS)
Tabla S1. Lista de los ortoflavivirus utilizados en este estudio.
https://doi.org/10.1371/journal.ppat.1014233.s003
(DOCX)
Tabla S2. Lista de secuencias SLA utilizadas con su estructura secundaria en notación de puntos y corchetes.
https://doi.org/10.1371/journal.ppat.1014233.s004
(DOCX)
Tabla S3. Diseños de PCR para la construcción de virus quiméricos.
https://doi.org/10.1371/journal.ppat.1014233.s005
(DOCX)
Tabla S4. Oligonucleótidos utilizados para PCR superpuestas para la construcción de virus quiméricos.
https://doi.org/10.1371/journal.ppat.1014233.s006
(DOCX)
Tabla S6. Secuencias de cebadores para la construcción de ARN SLA para ensayos de unión in vitro.
https://doi.org/10.1371/journal.ppat.1014233.s008
(DOCX)
Expresiones de gratitud
Los autores agradecen a los miembros del laboratorio Gamarnik por las discusiones y la revisión crítica del manuscrito. A Diego Álvarez y Eliana F. Castro por sus consejos con los experimentos de SM. También agradecemos a Christopher Laudeman por la síntesis de DMA-213, a TinTin Luu por la síntesis de DMA-227 y DMA-236, y a Alexander Pletnev y Sonja Best (NIAID/NIH) por proporcionar el virus Langat. Asimismo, agradecemos al centro de supercomputación de la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires, CeCAR.
Referencias
- 1.Pierson TC, Diamond MS. La continua amenaza de los flavivirus emergentes. Nat Microbiol. 2020;5(6):796–812. pmid:32367055
- 2.Guzmán MG, Harris E. Dengue. The Lancet. 2015;385:453–65.
- 3.Worden-Sapper ER, Gendler P, Lange RE, Kuhn JH, Sawyer SL. Amenazas futuras: ortoflavivirus animales que actualmente infectan a menos de 100 personas al año. Cell Rep. 2025;44(6):115700. pmid:40440170
- 4.Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. La distribución global y la carga del dengue. Naturaleza. 2013;496(7446):504–7. pmd:23563266
- 5.Dengue: situación mundial, vigilancia y progreso – Actualización 2024. https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue
- 6.Brasil P, Calvet GA, Siqueira AM, Wakimoto M, de Sequeira PC, Nobre A. Brote del virus Zika en Río de Janeiro, Brasil: caracterización clínica, aspectos epidemiológicos y virológicos. PLoS Negl Trop Dis. 2016;10:e0004636.
- 7.Monath TP, Vasconcelos PFC. Fiebre amarilla. J Clin Virol. 2015;64:160–73. pmid:25453327
- 8.Collins ND, Barrett ADT. Vacuna contra la fiebre amarilla 17D atenuada viva: una vacuna tradicional que aún controla brotes en la actualidad. Curr Infect Dis Rep. 2017;19.
- 9.Fischer M, Lindsey N, Staples JE, Hills S, Centros para el Control y la Prevención de Enfermedades (CDC). Vacunas contra la encefalitis japonesa: recomendaciones del Comité Asesor sobre Prácticas de Inmunización (ACIP). MMWR Recomm Rep. 2010;59(RR-1):1–27. pmid:20224546
- 10.Rendi-Wagner P. Avances en la vacunación contra la encefalitis transmitida por garrapatas. Expert Rev Vaccines. 2008;7(5):589–96. pmid:18564014
- 11.Tricou V, Yu D, Reynales H, Biswal S, Saez-Llorens X, Sirivichayakul C, et al. Eficacia y seguridad a largo plazo de una vacuna tetravalente contra el dengue (TAK-003): resultados a 4,5 años de un ensayo clínico de fase 3, aleatorizado, doble ciego y controlado con placebo. Lancet Glob Health. 2024;12(2):e257–70. pmid:38245116
- 12.Kallás EG, Cintra MAT, Moreira JA, Patiño EG, Braga PE, Tenório JCV. Vacuna viva atenuada tetravalente contra el dengue de butantán en niños y adultos. N Inglés J Med. 2024;390:397–408.
- 13.van den Elsen K, Chew BLA, Ho JS, Luo D. Proteínas no estructurales y complejos de replicación de flavivirus como dianas para fármacos antivirales. Curr Opin Virol. 2023;59:101305. pmid:36870091
- 14.Bifani AM, Chan KWK, Borrenberghs D, Tan MJA, Phoo WW, Watanabe S, et al. Terapéuticas para las infecciones flavivirales. Res. antivirus. 2023;210:105517. pmd:36592668
- 15.Parsons J, Castaldi MP, Dutta S, Dibrov SM, Wyles DL, Hermann T. Inhibición conformacional del ARN del sitio de entrada ribosómica interna del virus de la hepatitis C. Nat Chem Biol. 2009;5(11):823–5. pmid:19767736
- 16.Paulsen RB, Seth PP, Swayze EE, Griffey RH, Skalicky JJ, Cheatham TE 3rd, et al. Cambio estructural inducido por inhibidores en el ARN del dominio IIa del IRES del VHC. Proc Natl Acad Sci US A. 2010;107(16):7263–8. pmid:20360559
- 17.Dibrov SM, Ding K, Brunn ND, Parker MA, Bergdahl BM, Wyles DL, et al. La estructura de un dominio de ARN del virus de la hepatitis C en complejo con un inhibidor de la traducción revela un modo de unión similar al de los ribointerruptores. Proc Natl Acad Sci US A. 2012;109(14):5223–8. pmid:22431596
- 18.Connelly CM, Moon MH, Schneekloth JS Jr. El papel emergente del ARN como diana terapéutica para moléculas pequeñas. Cell Chem Biol. 2016;23(9):1077–90. pmid:27593111
- 19.Zafferani M, Hargrove AE. Dirigir moléculas pequeñas a estructuras terciarias y cuaternarias de ARN biológicamente relevantes. Cell Chem Biol. 2021;28(5):594–609. pmid:33823146
- 20.Welsch S, Miller S, Romero-Brey I, Merz A, Bleck CKE, Walther P, et al. Composición y arquitectura tridimensional de los sitios de replicación y ensamblaje del virus del dengue. Cell Host Microbe. 2009;5(4):365–75. pmid:19380115
- 21.Gillespie LK, Hoenen A, Morgan G, Mackenzie JM. El retículo endoplasmático proporciona la plataforma de membrana para la biogénesis del complejo de replicación del flavivirus. J Virol. 2010;84(20):10438–47. pmid:20686019
- 22.Ackermann M, Padmanabhan R. La síntesis de novo de ARN por la ARN polimerasa dependiente de ARN del virus del dengue muestra dependencia de la temperatura en la fase de iniciación, pero no en la de elongación. J Biol Chem. 2001;276(43):39926–37. pmid:11546770
- 23.Selisko B, Dutartre H, Guillemot JC, Debarnot C, Benarroch D, Khromykh A, et al. Estudios mecanísticos comparativos de la síntesis de ARN de novo por ARN polimerasas dependientes de ARN de flavivirus. Virology. 2006;351(1):145–58. pmid:16631221
- 24.Kao CC, Singh P, Ecker DJ. Iniciación de novo de la síntesis de ARN dependiente de ARN viral. Virología. 2001;287:251–60.
- 25.Zhao Y, Soh TS, Zheng J, Chan KWK, Phoo WW, Lee CC, et al. La estructura cristalina de la proteína NS5 del virus del dengue revela una nueva interfaz interdominio esencial para la flexibilidad de la proteína y la replicación viral. PLoS Pathog. 2015;11(3):e1004682. pmid:25775415
- 26.Yap TL, Xu T, Chen YL, Malet H, Egloff MP, Canard B. Estructura cristalina del dominio catalítico de la ARN polimerasa dependiente de ARN del virus del dengue a una resolución de 1,85 angstroms. J Virol. 2007;81:4753–65.
- 27.Egloff MP, Benarroch D, Selisko B, Romette JL, Canard B. Una metiltransferasa de (nucleósido-2′-O-)-cap de ARN en la ARN polimerasa NS5 del flavivirus: estructura cristalina y caracterización funcional. EMBO J. 2002;21:2757–68.
- 28.Zhou Y, Ray D, Zhao Y, Dong H, Ren S, Li Z. Estructura y función de la metiltransferasa NS5 del flavivirus. J Virol. 2007;81:3891–903.
- 29.Zhao Y, Soh TS, Lim SP, Chung KY, Swaminathan K, Vasudevan SG, et al. Base molecular para el reconocimiento específico del ARN viral y la metilación de la 2′-O-ribosa por la proteína no estructural 5 (NS5) del virus del dengue. Proc Natl Acad Sci US A. 2015;112(48):14834–9. pmid:26578813
- 30.Lindenbach BD, Thiel HJ, Rice CM. Flaviviridae: los virus y su replicación. Fields Virology. 2003. pp. 1101–52.
- 31.Filomatori CV, Lodeiro MF, Alvarez DE, Samsa MM, Pietrasanta L, Gamarnik AV. Un elemento de ARN 5′ promueve la síntesis de ARN del virus del dengue en un genoma circular. Desarrollo de genes. 2006;20(16):2238–49. pmid:16882970
- 32.Álvarez DE, Lodeiro MF, Ludueña SJ, Pietrasanta LI, Gamarnik AV. Las interacciones ARN-ARN de largo alcance circularizan el genoma del virus del dengue. J Virol. 2005;79(11):6631–43. pmid:15890901
- 33.You S, Padmanabhan R. Un nuevo sistema de replicación in vitro para el virus del dengue. J Biol Chem. 1999;274(47):33714–22.
- 34.Khromykh AA, Meka H, Guyatt KJ, Westaway EG. Papel esencial de las secuencias de ciclación en la replicación del ARN de los flavivirus. J Virol. 2001;75(14):6719–28. pmid:11413342
- 35.Zeng L, Falgout B, Markoff L. Identificación de secuencias de nucleótidos específicas dentro del 3′-SL conservado en el genoma del virus del dengue tipo 2, necesarias para la replicación. J Virol. 1998;72(9):7510–22. pmid:9696848
- 36.You S, Falgout B, Markoff L, Padmanabhan R. La síntesis de ARN in vitro a partir de plantillas de ARN viral del dengue exógeno requiere interacciones de largo alcance entre las regiones terminales 5′ y 3′ que influyen en la estructura del ARN. J Biol Chem. 2001;276(19):15581–91. pmid:11278787
- 37.Tilgner M, Shi PY. Estructura y función de los seis nucleótidos terminales 3′ del genoma del virus del Nilo Occidental en la replicación viral. J Virol. 2004;78(15):8159–71. pmid:15254187
- 38.Yu L, Markoff L. La topología de las protuberancias en el tallo largo del bucle del tallo 3′ del flavivirus es un determinante principal de la competencia de replicación del ARN. J Virol. 2005;79(4):2309–24. pmid:15681432
- 39.Khromykh AA, Kondratieva N, Sgro JY, Palmenberg A, Westaway EG. Importancia de los nucleótidos terminales del genoma del flavivirus en la replicación. J Virol. 2003;77(19):10623–9. pmid:12970446
- 40.Yu L, Nomaguchi M, Padmanabhan R, Markoff L. Requisitos específicos para los elementos de las regiones terminales 5′ y 3′ en la síntesis de ARN de flavivirus y la replicación viral. Virology. 2008;374(1):170–85. pmid:18234265
- 41.Bujalowski PJ, Bujalowski W, Choi KH. Identificación del sitio de unión del bucle del tallo A (SLA) del promotor del ARN viral en la polimerasa NS5 del virus Zika. Sci Rep. 2020;10(1):13306. pmid:32764551
- 42.Upstone L, Colley R, Harris M, Goonawardane N. Caracterización funcional de las estructuras secundarias del ARN de la región 5′ no traducida (UTR) en la replicación del virus de la encefalitis transmitida por garrapatas en células de mamíferos. PLoS Negl Trop Dis. 2023;17(1):e0011098. pmid:36689554
- 43.Lee E, Bujalowski PJ, Teramoto T, Gottipati K, Scott SD, Padmanabhan R, et al. Las estructuras de los promotores de ARN de flavivirus sugieren dos modos de unión con la polimerasa NS5. Comuna Nacional. 2021;12(1):2530. pmd:33953197
- 44.Sun YT, Varani G. Estructura del promotor del ARN del virus del dengue. RNA. 2022;28(9):1210–23. pmid:35750488
- 45.Osawa T, Aoki M, Ehara H, Sekine SI. Estructuras de complejos de ARN replicasa del virus del dengue. Celda Mol. 2023;83(15):2781–2791.e4. pmd:37478848
- 46.Lodeiro MF, Filomatori CV, Gamarnik AV. Estudios estructurales y funcionales del elemento promotor de la replicación del ARN del virus del dengue. J Virol. 2009;83(2):993–1008. pmmid:19004935
- 47.Dethoff EA, Boerneke MA, Gokhale NS, Muhire BM, Martin DP, Sacco MT, et al. Estructura terciaria generalizada en el genoma de ARN del virus del dengue. Proc Natl Acad Sci US A. 2018;115(45):11513–8. pmid:30341219
- 48.Huber RG, Lim XN, Ng WC, Sim AYL, Poh HX, Shen Y, et al. El mapeo estructural de los virus del dengue y del Zika revela interacciones funcionales de largo alcance. Nat Commun. 2019;10(1):1408. pmid:30926818
- 49.Obi JO, Kihn KC, McQueen L, Fields JK, Snyder GA, Deredge DJ. Dinámica estructural de las interacciones de la proteína no estructural 5 (NS5) del virus del dengue con el bucle del tallo A (SLA) del promotor. Npj Viruses. 2025;3(1):30. pmid:40295851
- 50.Pallarés HM, Costa Navarro GS, Villordo SM, Merwaiss F, de Borba L, Gonzalez Lopez Ledesma MM, et al. La generación de ARN subgenómico del flavivirus del virus Zika requiere cooperación entre estructuras de ARN duplicadas que son esenciales para la infección productiva en células humanas. J Virol. 2020;94(18):e00343–20. pmid:32581095
- 51.de Borba L, Villordo SM, Iglesias NG, Filomatori CV, Gebhard LG, Gamarnik AV. Las interacciones ARN-ARN locales y de largo alcance superpuestas modulan la ciclización y replicación del genoma del virus del dengue. J Virol. 2015;89(6):3430–7. pmid:25589642
- 52.Selisko B, Potisopon S, Agred R, Priet S, Varlet I, Thillier Y, et al. Base molecular de la conservación de nucleótidos en los extremos del genoma del virus del dengue. PLoS Pathog. 2012;8(9):e1002912. pmid:23028313
- 53.Dong H, Ray D, Ren S, Zhang B, Puig-Basagoiti F, Takagi Y, et al. Los distintos elementos de ARN confieren especificidad a los eventos de metilación de la tapa de ARN de flavivirus. J Virol. 2007;81(9):4412–21. pmd:17301144
- 54.Feinberg MA, Le MT, Carpio KL, Knyazhanskaya E, Barrett ADT, Choi KH. Interacción de la proteína NS5 del virus del Nilo Occidental con los ARN SLA de ortoflavivirus y sus efectos sobre la replicación e inhibición viral. J Virol. 2025;99(9):e0202324. pmid:40824093
- 55.Wang S, Chan KWK, Tan MJA, Flory C, Luo D, Lescar J, et al. Una arginina conservada en NS5 se une al ARN genómico con bucle en tallo 3′ para la iniciación independiente del cebador de la replicación del ARN del flavivirus. RNA. 2022;28(2):177–93. pmid:34759006
- 56.Lescoute A, Westhof E. Topología de las uniones de tres vías en ARN plegados. RNA. 2006;12(1):83–93. pmid:16373494
- 57.Keating KS, Humphris EL, Pyle AM. Una nueva forma de ver el ARN. Q Rev Biophys. 2011;44:433–66.
- 58.Waskom M. seaborn: visualización de datos estadísticos. J Open Source Softw. 2021;6(60):3021.
- 59.Abram QH, Matthews LA, Guarné A, Sagan SM. Caracterización estructural y funcional de la estructura SLA’ en el extremo 3′ del intermediario de cadena negativa del virus Zika. RNA. 2025;31(8):1139–53. pmid:40341208
- 60.Zafferani M, Martyr JG, Muralidharan D, Montalvan NI, Cai Z, Hargrove AE. El perfilado multiensayo de una biblioteca de moléculas pequeñas focalizada revela una modulación bidireccional predictiva de la estabilidad del triplex del lncRNA MALAT1 in vitro. ACS Chem Biol. 2022;17(9):2437–47. pmid:35984959
- 61.Patwardhan NN, Cai Z, Umuhire Juru A, Hargrove AE. Factores determinantes en el reconocimiento de dianas de ARN del VIH por amilorida. Org Biomol Chem. 2019;17(42):9313–20. pmid:31612165
- 62.Wicks SL, Morgan BS, Wilson AW, Hargrove AE. Explorando el espacio químico bioactivo para descubrir moléculas pequeñas dirigidas al ARN. 2023.
- 63.Campbell MS, Pletnev AG. Clones de ADNc infecciosos del flavivirus transmitido por garrapatas de Langat que difieren de su progenitor en la neurovirulencia periférica. Virology. 2000;269(1):225–37. pmid:10725214
- 64.Umuhire Juru A, Hargrove AE. Estructuras para la focalización de ARN con moléculas pequeñas. J Biol Chem. 2021;296:100191. pmid:33334887
- 65.Zafferani M, Haddad C, Luo L, Dávila-Calderón J, Chiu LY, Mugisha CS, et al. Las amiloridas inhiben la replicación del SARS-CoV-2 in vitro al atacar las estructuras del ARN. Ciencia avanzada. 2021;7(48):eabl6096. pmd:34826236
- 66.Patwardhan NN, Ganser LR, Kapral GJ, Eubanks CS, Lee J, Sathyamoorthy B, et al. Amilorida como un nuevo andamio de unión a ARN con actividad contra el TAR del VIH-1. Medchemcomm. 2017;8(5):1022–36. pmid:28798862
- 67.Davila-Calderon J, Patwardhan NN, Chiu LY, Sugarman A, Cai Z, Penutmutchu SR, et al. Una molécula pequeña dirigida al IRES inhibe la replicación del enterovirus 71 mediante la estabilización alostérica de un complejo ternario. Nat Commun. 2020;11(1):4775. pmid:32963221
- 68.Davila-Calderon J, Li ML, Penumutchu SR, Haddad C, Malcolm L, King J, et al. La evolución del enterovirus revela el mecanismo de un antiviral dirigido al ARN y los determinantes de la replicación viral. Sci Adv. 2024;10(7):eadg3060. pmid:38363831
- 69.Tangudu CS, Dankaona W, Blitvich BJ. El intercambio del promotor A con estructura de tallo-bucle del virus Zika por la región correspondiente del virus de la encefalitis transmitida por garrapatas genera un virus quimérico que se replica en células de vertebrados y mosquitos. Arch Virol. 2025;170(12):236. pmid:41162790
- 70.Brillet K, Janczuk-Richter M, Poon A, Laukart-Bradley J, Ennifar E, Lebars I. Caracterización del promotor de ARN SLA del virus del dengue y su interacción con la proteína no estructural viral NS5. Biochimie. 2024;222:87–100. pmid:38408720
- 71.El Sahili A, Soh TS, Schiltz J, Gharbi-Ayachi A, Seh CC, Shi PY, et al. NS5 del serotipo 2 del virus del dengue puede adoptar una conformación análoga a la de sus homólogos del virus Zika y del virus de la encefalitis japonesa. J Virol. 2019;94(1):e01294–19. pmd:31597763
- 72.Potisopon S, Priet S, Collet A, Decroly E, Canard B, Selisko B. El dominio metiltransferasa de la proteína NS5 del virus del dengue garantiza la iniciación y elongación eficientes de la síntesis de ARN por el dominio polimerasa. Nucleic Acids Res. 2014;42(18):11642–56. pmid:25209234
- 73.Best SM. Las múltiples facetas de la proteína NS5 del flavivirus en el antagonismo de la señalización del interferón tipo I. J Virol. 2017;91(3):e01970–16. pmid:27881649
- 74.Ashour J, Laurent-Rolle M, Shi PY, García-Sastre A. NS5 del virus del dengue media la unión y degradación de STAT2. J Virol. 2009;83(11):5408–18. pmmid:19279106
- 75.González López Ledesma MM, Costa Navarro G, Pallares HM, Paletta A, De Maio F, Iglesias NG, et al. El virus NS5 del dengue degrada ERC1 durante la infección para antagonizar la activación de NF-kB. Proc Natl Acad Sci US A. 2023;120(23):e2220005120. pmd:37252973
- 76.Sievers F, Higgins DG. Clustal Omega para realizar alineamientos precisos de múltiples secuencias de proteínas. Protein Sci. 2018;27(1):135–45. pmid:28884485
- 77.Edgar RC. MUSCLE: alineación de secuencias múltiples con alta precisión y alto rendimiento. Nucleic Acids Res. 2004;32(5):1792–7. pmid:15034147
- 78.Crooks GE, Hon G, Chandonia JM, Brenner SE. WebLogo: un generador de logotipos de secuencias: Figura 1. Genome Res. 2004;14:1188–90.
- 79.Trifinopoulos J, Nguyen LT, von Haeseler A, Minh BQ. W-IQ-TREE: una herramienta filogenética rápida en línea para el análisis de máxima verosimilitud. Nucleic Acids Res. 2016;44(W1):W232–5. pmid:27084950
- 80.Letunic I, Bork P. Interactive Tree of Life (iTOL) v6: actualizaciones recientes de la herramienta de visualización y anotación de árboles filogenéticos. Nucleic Acids Res. 2024;52(W1):W78–82. pmid:38613393
- 81.Huston NC, Tsao LH, Brackney DE, Pyle AM. El genoma del virus del Nilo Occidental alberga elementos riborreguladores esenciales con funciones conservadas y específicas del huésped. Proc Natl Acad Sci US A. 2024;121(29):e2312080121. pmid:38985757
- 82.Li P, Wei Y, Mei M, Tang L, Sun L, Huang W, et al. Análisis integrador de la estructura del ARN del genoma del virus Zika revela determinantes críticos de la infectividad viral. Cell Host Microbe. 2018;24(6):875–886.e5. pmid:30472207
- 83.Li D, Lu HT, Ding YZ, Wang HJ, Ye JL, Qin CF, et al. Elementos de ARN cis-activos especializados equilibran la ciclación del genoma para asegurar la replicación eficiente del virus de la fiebre amarilla. J Virol. 2023;97(4):e0194922. pmid:37017533
- 84.Gruber AR, Lorenz R, Bernhart SH, Neuböck R, Hofacker IL. El conjunto de herramientas web de ARN de Viena. Nucleic Acids Res. 2008;36:W70–4. pmid:18424795
- 85.Mathews DH, Sabina J, Zuker M, Turner DH. La dependencia de secuencia ampliada de los parámetros termodinámicos mejora la predicción de la estructura secundaria del ARN. J Mol Biol. 1999;288(5):911–40. pmid:10329189
- 86.Kerpedjiev P, Hammer S, Hofacker IL. Forna (ARN dirigido por fuerza): Diagramas de estructura secundaria de ARN sencillos y eficaces en línea. Bioinformática. 2015;31(20):3377–9. pmid:26099263
- 87.Will S, Reiche K, Hofacker IL, Stadler PF, Backofen R. Inferencia de familias y clases de ARN no codificante mediante agrupamiento basado en la estructura a escala genómica. PLoS Comput Biol. 2007;3(4):e65. pmid:17432929
- 88.Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: modelado por homología de estructuras y complejos proteicos. Nucleic Acids Res. 2018;46(W1):W296–303. pmid:29788355
- 89.Rother M, Rother K, Puton T, Bujnicki JM. ModeRNA: una herramienta para el modelado comparativo de la estructura 3D del ARN. Nucleic Acids Res. 2011;39(10):4007–22. pmid:21300639
- 90.Iglesias NG, Filomatori CV, Gamarnik AV. El motivo F1 de la polimerasa NS5 del virus del dengue participa en la síntesis de ARN dependiente del promotor. J Virol. 2011;85(12):5745–56. pmid:21471248
- 91.Maier JA, Martinez C, Kasavajhala K, Wickstrom L, Hauser KE, Simmerling C. ff14SB: mejora de la precisión de los parámetros de la cadena lateral y el esqueleto proteico a partir de ff99SB. J Chem Theory Comput. 2015;11(8):3696–713. pmid:26574453
- 92.Aytenfisu AH, Spasic A, Grossfield A, Stern HA, Mathews DH. Los parámetros diedros del ARN revisados para el campo de fuerza Amber mejoran la dinámica molecular del ARN. J Chem Theory Comput. 2017;13(2):900–15. pmid:28048939
- 93.Peters MB, Yang Y, Wang B, Füsti-Molnár L, Weaver MN, Merz KM Jr. Estudio estructural de proteínas que contienen zinc y desarrollo del campo de fuerza Zinc AMBER (ZAFF). J Chem Theory Comput. 2010;6(9):2935–47. pmid:20856692
- 94.Li P, Merz KM Jr. MCPB.py: un generador de parámetros de centros metálicos basado en Python. J Chem Inf Model. 2016;56(4):599–604. pmid:26913476
- 95.Olsson MHM, Søndergaard CR, Rostkowski M, Jensen JH. PROPKA3: tratamiento consistente de residuos internos y superficiales en predicciones empíricas de pKa. J Chem Theory Comput. 2011;7(2):525–37. pmid:26596171
- 96.Caso DA, Aktulga HM, Belfon K, Cerutti DS, Cisneros GA, Cruzeiro VWD, et al. Herramientas ámbar. Modelo J Chem Inf. 2023;63:6183–91.
- 97.Jorgensen WL, Chandrasekhar J, Madura JD, Impey RW, Klein ML. Comparación de funciones potenciales simples para simular agua líquida. J Chem Phys. 1983;79:926–35.
- 98.Pastor RW, Brooks BR, Szabo A. Un análisis de la precisión de los algoritmos de Langevin y de dinámica molecular. Mol Phys. 1988;65:1409–19.
- 99.Berendsen HJC, Postma JPM, Van Gunsteren WF, Dinola A, Haak JR. Dinámica molecular con acoplamiento a un baño externo. J Química Física. 1984;81:3684–90.
- 100.Ryckaert JP, Ciccotti G, Berendsen HJC. Integración numérica de las ecuaciones cartesianas de movimiento de un sistema con restricciones: dinámica molecular de n-alcanos. J Comput Phys. 1977;23:327–41.
- 101.Sarzynska J, Popenda M, Antczak M, Szachniuk M. Predicción de la estructura terciaria del ARN mediante RNAComposer en CASP15. Proteins. 2023;91(12):1790–9. pmid:37615316
- 102.Humphrey W, Dalke A, Schulten K. VMD: dinámica molecular visual. J Mol Graph. 1996;14(1):33–8, 27–8. pmid:8744570
- 103.Pettersen EF, Goddard TD, Huang CC, Meng EC, Couch GS, Croll TI, et al. UCSF ChimeraX: Visualización de estructuras para investigadores, educadores y desarrolladores. Protein Sci. 2021;30(1):70–82. pmid:32881101
- 104.Lu XJ, Bussemaker HJ, Olson WK. DSSR: una herramienta de software integrada para analizar la estructura espacial del ARN. Nucleic Acids Res. 2015;43(21):e142. pmid:26184874
- 105.Kinney RM, Butrapet S, Chang GJJ, Tsuchiya KR, Roehrig JT, Bhamarapravati N. Construcción de clones de ADNc infecciosos para el virus del dengue 2: cepa 16681 y su derivado vacunal atenuado, cepa PDK-53. Virology. 1997;230:300–8.
- 106.Zhang J, Chung T, Oldenburg K. Un parámetro estadístico simple para la evaluación y validación de ensayos de cribado de alto rendimiento. J Biomol Screen. 1999;4(2):67–73. pmid:10838414
- 107.Bredenbeek PJ, Kooi EA, Lindenbach B, Huijkman N, Rice CM, Spaan WJM. Un clon estable de ADNc de longitud completa del virus de la fiebre amarilla y el papel de los elementos de ARN conservados en la replicación de los flavivirus. J Gen Virol. 2003;84(Pt 5):1261–8. pmid:12692292









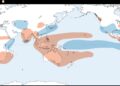



{kind=link}